Rauber-Kopsch解剖学
歴史的な偉大な解剖学書
Rauber Kopsch Band1. 04 


III. 形態上の要素について,すなわち細胞学Cytologia, Zellenlehre
A.慨説Allgemeinesおよび形態学的なことMorphologisches
動物と植物は細胞Cellula, Zelleとよばれるところの,多くは顕微鏡の力をかりてやっと見ることができ,何れも相似た性質をもつところの基本単位から構成されている.
定義:細胞は生活物質の1つの塊であって,細胞核Zellkernと細胞体Zelleibとよりなり,この塊は形態の上からも機能の上からも生物体の基本的な単位とみなされるのである.
成長を遂げた生物体では同一の組織内の細胞は多くの場合たがいに数多くの突起で結合している.もっともこの突起の数が少ないこともある.また異なる組織に属する細胞相互のあいだにも融合があって,かくして高度の機能的全一性がえられる(例えば神経細胞と筋細胞もしくは上皮細胞とのあいだ).しかしこの事実は独立した単位であるという性質を個々の細胞からうばいさるものではない.(Heidenhain, Plasma und Zelle.1907-Boeke, Med. Klinik.1925.-Patzelt, Z. mikr.-anat. Forsch., 3. Bd.,1925)
細胞がもつ高い一般的な意義は次に述べる概括からわかるのである:
1. 植物および動物のからは細胞あるいは細胞のつくったものからできている;
2. 植物および動物の体は細胞から出発する.その細胞は卵細胞と精細胞とである;
3. 細胞は生活機能を担うものである.すなわち生命の単位Lebenseinheitである(ウィルヒョウR. Virchowの細胞病理学Cellularpathologie);
4. 生物の系統において最も低い位置にあるものは単細胞の生物(Unicellulaten, Protisten)である.それより高等な生物の体は細胞が集まってそれをなしている.すなわち多細胞生物(Multicellulaten)である.これに後生殖物Metaphytenと後生動物Metazoenとがある.
おのおのの細胞は細胞体Zelleibと核Zellkernとからなる.
細胞膜Zellmembranは細胞の本質的なものに属しない.植物の細胞(図1)ではじようぶなセルローズの膜で囲まれているから細胞膜という名前が適当であるが,動物の細胞では特別な膜を探し求めても多くのばあいむだである.そこでは普通は細胞体の最も表層のところが密になっているにすぎない.
むかしHermann Lotze(Allgem. Physiol. d. Körperlichen Lebens, Leipzig,1951)が疑問としたことが今日でもやはり大きい疑問としてあらわれてきている.それはわれわれの知る限りのあらゆる生命が形の上で細胞と結びついているか,あるいはその他の形で起こりうるのではないかということである.われわれの知るすべての生命が原形質と核物質に結ばれているということができる.最も小さい生物diekleinsten Lebewesenである細菌でさえも細胞体の性質を示す部分と核の性質を示す部分,つまり2つの成分をもつのである.だからわれわれは細菌を特別な種類の細胞(最小細胞Minimo-Cellulae)と考えなければならない.
これに反して瀘過性病原体すなわちウィルスの各種は人,その他の動物や植物でたくさんの数(100以上)の伝染病(例えば天然痘,狂犬病,ハシカ)をひきおこすものであるが,もはや細胞ではないかも知れない.何となれば最も小さいウィルスは1個のタンパク質分子よりも大きくはない.それは細胞になる前の有機体であって,その最も簡単な種類のものは核タンパク質からのみできていて,複雑な種類のウィルスはその他にリポイドと含水炭素を含んでおり,その組成からして細菌と違っていないものである.
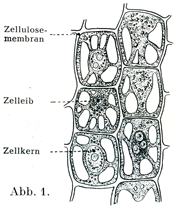
[図1]植物細胞(Sachsによる.)
細胞の原形Grundformは球形である.しかし後生動物の体をなしている細胞の多くはその他の形を示している.楕円に近いもの,錐体すなわちピラミッドの形のもの,柱状,円板状,星状のものがあり,その各々にも千差万別があるといえるのである.
大きさGrößeは平均すると約20µである.血球やリンパ性の細胞では2~6µのものまであり,微生物のあるものはもっとそれより小さい,他の細胞ではひじょうに大きくなっていて,例えば魚類,両棲類,鳥類の卵のように,空間の三次元において何れの方向にも大きいものがあり,あるいは神経細胞のように,特に長さだけがはなはだ延びていることがある.神経細胞はその軸索突起をもって脊髄から手や足の指まで達しているので,その長さが1メートルもある.
いろいろな種類の細胞について,細胞体とかくの体積の比をしらべてみると,たがいに一定した値になる.これを核と形質の比価Kern-Plasma-Relationという.
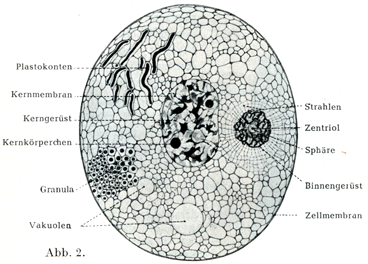
[図2]原形質の蜂巣状構造を示す細胞模型図
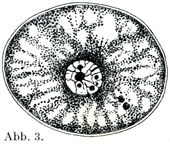
[図3]コウモリ卵巣にて成熟しつつある卵.原形質は小梁状の構造をしっめし,そのあいだに液胞をもっている.切片標本による.×330. (Rauber, Morph. Jahrb. VIII.,1883, Neue Grundlegungen usw. より.)
I. 細胞体Zelleb
化学的および物理的組成:細胞体は原形質よりなる.
原形質Protoplasmあるいは細胞形質Cytoplasmaというのは化学的に単一な物質の名前ではなくて,1つの形態学的な概念である.それはいくつかの蛋白体と(とくにプラスチンPlastinならびにグロブリンglobuline, アルブモーゼAlbumosen,アルブミンAlbumine),多量の水分および数種の塩基からなっている.そのほかに同化性および異化性の物質代謝産物が加わる.反応はアルカリ性であって,みな一様で,粘い液のような性質をもっている.水によって膨化し,熱によって凝固する.
構造:だいじな問題としては原形質ではその各成分が細胞のなかで溶液の形のみ存在し,つまりその各々が密に混合した状態なのか,もしくは原形質に何か特別の構造があるかということである.むしろその構造がどんな種類であるかについて議論が分かれている.すなわち海綿様の構造であるというもの(Fromman, Schmitz, Leydig),蜂巣状となすもの(Bütschli),あるいは細かい糸の構造とその間を満たす物質からなるという人がある(Flemming).
この3つの学説のほかになおAltmannの独特な顆粒説がある.その何れが正しいかを決定するのはその研究の困難からしてはなはだむつかしいことである.しかし,蜂巣説の考え方が次第に大きい力を得つつある.その一員は生理化学的の事実がこの説に都合よくあてはまることである.
Bütschliによれば原形質の構造は泡様schaumigであって,蜂の巣を連想させるものであるという.しかし,この説によっては原形質の構造に関する問題のほんの一部が答えられているにすぎない.蜂の巣の壁をなす索や板がどんなこうぞうであるかについて問題がまた戻ってくるのである,そいう超構造Ultrastrukturは比較的近年にやっと研究されてきた.(Bargmannの著Lehrbuch der Histologie, Bd. Iを参照されたい.)
顆粒は原形質内の構造として常にみられるものである.これは生きている細胞において(Ehrlich , Arnold, Fischel)メチレン青,中性赤そのほかの数多くのいわゆる生体染色用の色素によってあらわすことができる.しかしまた,顆粒にもいろいろの種類が区別できる.顆粒は総括して微小粒体Mikrosomen,さらによい名前では形質粒体Plasmosomenとよばれる.
生体染色によってあらわされる顆粒と区別すべきものは固定剤のためにアルブモーゼが沈殿することにより人工的にできた顆粒である(A. Fischer, Fixierung usw. des Protoplasmas. Jena,1899).Altmannが記載した顆粒の一部は生きている細胞には存在せずして,顕微鏡でしらべる目的で細胞を薬品で固定したときに人工的につくられたものであることが確かである.
微小粒体が相ならんで糸の形をしているのを最初Brunnが1884年,La Valette Sr. Georgeが1886年に記載し,それが後にミトコンドリア(糸粒体)MitochondrienあるいはChondriosomenとよばれることになった.このものは成体および胎児の体の多くの細胞に証明されている(Duesbergの文献をみられたい).Mevesはミトコンドリアをあらわすのに適当した方法を用いたときにつよく染まるが,個々の顆粒からなることがはっきりしないような糸状および棒状のものを粒杆体Chondriokonten(κουtός,棒のこと)とよんだ(1907).その後にMevesはこれらの構造物を細胞に独特な成分として大いに重要視し,原形質の遺伝を担当するものとみなして,形成顆粒Plastosomenと名づけた.その顆粒状のものをPlastokonten(形成杆体)とよんだ.それより前に用いたChondriokontenやChondriosomenという名称はやめたのである.Mevesは次のごとく述べている(Meves, Arch. mikr. Anat.,85. Bd.,1914, S. 298).“形成粒体は独特な性質の顆粒あるいは糸状のものであって,すでに生きている細胞でたくさんに見ることができる.
それは胎児ではすべての細胞に存在し,成体でも数多くの細胞に証明される”.
Duesberg, J., Merlel und Bonnet, Ergebnisse, 20. Bd.1912.-meves, Arch. mikr. Anat.,94. Bd.,1920.
細胞体のもつ特殊な器官としては1. 中心小体とその周りの中心球,2. 内網装置,3. 分泌細管および分泌液胞である.
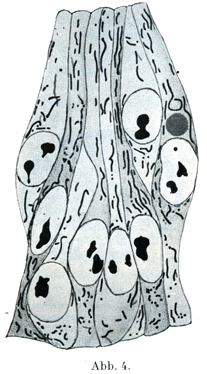
[図4]形成杆体Plastokonten. 孵化27時間(8対の原節を有する)ニワトリの胎児の神経管Medullarrohrの外胚葉細胞群.

[図5]形成粒体Plastochondrien.1個の内胚葉細胞.
1. 中心球Zentrosphareと中心小体Zentrolkorperchen
中心球は多くは円い形をした細胞小器官であって,原形質のなかにあるが,もともとはおそらく細胞核からできたものであろう.これは色素に対して特別な反応を示す細胞形質の1塊であって,旧形質Archoplasma (Boveri)とよばれ,その中にやっと認められるほどの小さい,そして光をつよく屈折する粒子があって,これが中心小体Zentrolkrörperchen (Flemming,1891)あるいは中心子Centriolum (Boveri,1895)とよばれる.中心子が1つだけでなく,それが2つあるいはそれ以上もあることがはなはだしばしばである(図8,9,10).中心小体のすぐまわりをとりまいて明るい幅のせまい部分があり,その外側に暗くて顆粒に富む.比較的に幅の広い部分があって,これが細胞体の残りの部分に直接つづくのである.このくらい部分が球Sphäreとよばれる(図6).中心球から周囲にむかって星Asterとよばれるつよい放射状のすじがでている.このすじが上部の暗い部分を通って細胞体のなかにつづいている(図7).中心球はそれ自身の分裂のほかに細胞核の分裂や細胞体の分裂を開始させるという大事な役目をもっている.つまり細胞分裂のための器官である.
それ故に中心小体はまた“運動の中心”Kinozentrumともいわれる.これに対して核は“物質代謝の中心”Chemozentrumということができる.しかし留意すべきことは,中心小体がなくても原形質の運動は起こりえることである.それは切りはなされた偽足が,かなり長い間運動を続けることによって示される.
中心小体は細胞分裂にあたって初めて出現するのではなくて,分裂以外の休止期にも存在する.休止期の細胞に中心小体が見られるものの例とそして白血球(図10,16),上皮細胞,結合組織細胞(図8,9)をあげておこう.しばしば核の縁にへこみがあって, そこに中心小体が存在している.
中心子の数と形は変化に富んでいる.動物の種類や組織の種類の違いによって2つあるいは3つの多くは円い形の中心子がみられる.2つのときは双心子Diplosomenという.数はおそらく時間的にもちがうのであろう.中心小棒Zentralstäbchenというのは細長くのびた棒状の中心子であって,最初にZimmermannによって或る硬骨魚の黄色い色素細胞においてみつけられて,その後ほかの学者たちによっていろいろな動物の雄の生殖細胞でもみられたのである.
中心球あるいは中心子の何れにいっそう重要な形態学上および生理学上の意味があるかは,なお決定していない.Mevesおよびその他の人びとは中心小体すなわち中心子の存在に重きを置いている.その理由は組織細胞分裂にあたって,中心小体はよく見えるが,球は多くの場合に存在しないことである.
歴史:Edurd van Benedenが1876年に初めて二杯虫類Dicyemidenの卵において中心球を発見し,ついで1883年に蛔虫Ascarisにおいてみた.-Heidenhain, M., Plasma und Zelle. Jena,1907.-Henneguy, La Cellule.-Hertwig, O., Die Zelle. Jena. G. Fischer.-Wilson, H. V., The Cell.-Mollendorf, W. von, Handbuch der mikr. Anat. Berlin,1926~52. )
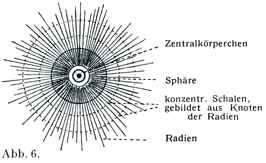
[図6]中心球の構造を示す模型図
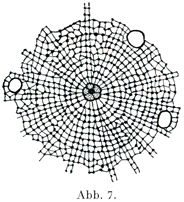
[図7]中心球と星の微細構造(Erlanger,1897による模型図.)
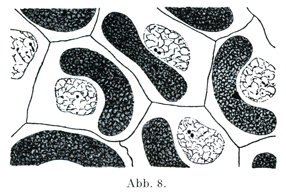
[図8]1つないし3つの中心子をもつ中心球3~4才のネコの角膜の内境界膜に接する内皮細胞.(E. Ballowitz,1900)

[図9]2つの中心子をもつZentrophormiumネコの前眼房の内皮細胞1個.(Ballowitz,1900)
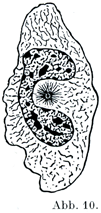
[図10]サンショウウォの幼生の腹膜よりえられた白血球.中心小体は明るい部分に取り囲まれて,更にその周囲に放射状の球Sphäreがある.(W. Flemming)
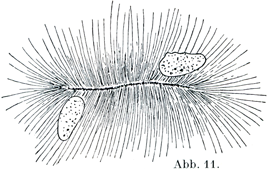
[図11]中心小棒Zentralstab,すなわち棒状をなす中心小体,と2個の核.硬骨魚Sargus(棘鰭類に属する)の黄色の色素塊にてみられたもの.(K. W. Zimmermann)
2. 内網装置Binnengerust, Endopegma 1)
1)細胞内の足場(建築のときに組み立てる)の意.(小川鼎三)
内網装置は前にはBinnennetz (Kopsch,1902)とよれば,いまでは多くのばあいゴルジ装置Golgi-Apparatとよばれるが,Binnengerüst, Endopegma (Kopsch,1925)というのが最もよいと考えられる.これは脊椎および無脊椎動物のすべての細胞に存在する細胞器官である.
以前に“副核”Nebenkernとよばれたものは内毛瘡土地,細胞体の一部でその網のあいだにある部分を合わせたもので,ときには中心球もそれに属している.
形態:内網装置は成長した脊椎動物の細胞では多くの場合に最大いろいろの円柱状の小梁を組み合わせた形であって,それが糸毬状をなし(図12),あるいは中空の球の壁をなしている(図9,13).
位置:上皮細胞ではそれは多くは核と細胞の自由面とのあいだにある.すなわち核の上方oberhalbdes Kernsにある(図12).しかしときとして核と細胞底とのあいだ,すなわち核の下方unterhalb des Kernsにある.まれには核のまわりをかこんでいる.神経細胞では概してそうである(図124).平たい薄い細胞では内網装置の大部分がふつうに核のそばneben dem Kernにある(図8,9).
中心球との関係:内皮細胞(図9)および生殖細胞(図13)では内網装置は中心球を囲んで中空の球の壁のようになっている.ほかの種類の細胞でも(とくに上皮細胞では)両者の同様な位置関係があるらしい.これを単に偶然とみなすべきであるのか,そして全く局所解剖学的に論ずるべきものであるのか,あるいは内網装置と中心球とが機能のうえで,またはその他の何らかの都合でたがいに結びついているものなかは現在のところ分かっていない.
前眼房の内皮細胞ではBallowitzが両者の関係からしてZentrophormiumと名づけ,生殖細胞ではHeidennhainが全体としてZentralkapsel(中心球の嚢の意か),その個々の成分をPseudochromosomen(偽染色体の意)とよんだ.最後に述べたものはHermannのArchoplasmaschleifen(旧形質係蹄)に相当し,これはすでに1885年にPlatnerによってNebenkernstäbchen(副核小棒)およびNebenkernfäden)として記載さえたものである.
意義:はなはだ多数の研究がなされたに関わらず,内網装置の意義については今でもなお確たることが何も言えないのである.確信できるのはこの装置をなしている物質の量と細胞の活動力とのあいだに連関があって,能動性の細胞は受動性のものよりいっそう大きい内網装置をもつ(Kopsch,1926, S. 276)ということだけである.また内網装置が精子の前端部のいわゆるAkrosomの形成に関与している事実ははなはだ重要なことにちがいないが,今でまだ充分にその価置が認識されていない.
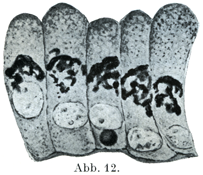
[図12]44才の男の前立腺の円柱細胞における糸毬状の内網装置.×1500. Kopsh-Kolatschevの方法. (Kopsch,1926)
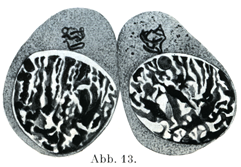
[図13]プロテウスProteus(有尾両棲類)の精細胞における偽染色体Pseudochromosomen (Heidenhain,1900)
細胞分裂のときの内網装置:核分裂および細胞分裂にあたって内網措置はばらばらのものになる.これをPegmatosomen (Kopsch,1925),またはDiktyosomen (Prroncito,1910)という.これらは2つの娘細胞に分配される(もっとも平等な分配というわけでないが),そして娘細胞のなかでふたたびまとまって内網装置ができる.
化学的組成:内網装置の化学的組成は動物の種類によって違い,同一の個体でも細胞の種類によって異なり,さらに胎児と成長した動物とのあいだでもちがうのである.このものはオスミウム酸を還元するといわゆる好性物質osmiophile Substanzとオスミウム酸を還元しないいわゆるオスミウム嫌性物質osmiophobe Substanzとから成っている.前者は酸性の保存液のなかではなくなってしまう.しかしこの両物質がいかなる種類のものであるかは今日まだはっきりしていない.
酸性の保存液を用いた場合に内網装置の梁のかわりに細い管があらわれる.これがHolmgrenのいう液細管Saftkanälchenである.また特別な染め方でオスミウム嫌性物質をそめて現わしたものが,Holmgrenの栄養海綿体Trophospongiumである.
歴史:すでに1885年にPlatnerが記述し,画にもかいたNebenkernstäbchen(副核小棒)およびNebenkernfäden(副核糸)は内網装置の一部が不完全にあらわれたものであった.Hermann(1891)のArchoplasmaschleifen(旧形質係蹄)およびHeidenhain(1900)のZentralkapseln(中心球の嚢)およびPseudochromosomen(偽染色体)についても同様のことが云える.内網装置が完全な形であらわされたのはGolgi(1898)が最初であって,GolgihaはそれをApparato reticolare interno(内網装置)とよび,またCntrophormiumという名称をつけたBallowitzも最初の開拓者の1人である.
オスミウム酸によって(Kopsch,1902)あらわされた内網装置はKopsch自身によってGolgiがすでに発表していたものと同一のものとみなされた.しかしのちにいっそう適切な名前としてBinnengerüst, Endopegma(ずれも細胞内部の足場の意)が提唱されたのである(Kopsh, Fr. : Enzylkopädlie mikr. Technik, 3. Aufl.,1926, Bd.1, S.163).
3. 分泌細管Sekretkapillarenと分泌液胞Sekretvakuolen
腺細胞のもつ分泌液胞は多くは円い形の小さい空所であって,分泌物あるいは排出物をもってみたし,その内容は腺細胞の自由面すなわち腺腔にむかっている表面に出されるのである.
細胞内分泌細管binnenzellige Sekretkapillarenは腺細胞の原形質のなかにある不分枝の,あるいは分枝した,ときとしては網状になっている管であって,腺細胞の自由面に開口している.(本書第II巻胃底腺の傍細胞を参照のこと.)
特別の細胞膜Zellmembran, すなわち原形質とははっきりと境された一つとつづきの膜状のものが細胞体の縁をとりまくことは多くの動物細胞にはみられないことは前述のごとくである.その代わりに原形質の最も表層のところが密になった1層をしている.はっきりと区別できる細胞膜は卵細胞に存在する.脂肪細胞や横紋筋線維にもあるが,この2者では細胞膜は周囲の結合組織がつくったものといわれている.
II. 細胞核Nucleus, Zellkern
定義:細胞核は特殊の化学物質(核物質Kernsubstanzen)より成り,細胞体とは異なる組成であって後者との境はある程度はっきりしおり,また細胞内で核の形はいろいろ異なる様子をしめすのである(Hertwig, O., Zelle).
核をなす物質は核形質Karyoplasma (κaρυον=核)とよばれて,細胞体をなす細胞形質Cytoplasmaと対応さえる.
核の形ははなはだ多様である.多くは球形(図14,15)であり,また卵円あるいはレンズ形であるが,なお乳頭状のもの,膨れだした形のもの,腎臓形のもの(図8~10),分葉状のもの(図16),分枝するものはおよび珠数状のものがある.
位置:多くのばあい,細胞中心に近く存在するが,またある程度の差はあるが中心を外れていることがしばしばあり,またその存在する場所を変えることができるものである.
核の大きさは大体に細胞の大きさにある程度伴っている.平均して4~9µである.人体で最大の核といえば卵細胞のもの(45µ)と,脊髄神経節の細胞のも能登である.
Jakobj(Verh. anat. Ges.,1931)は人体で,同一の器官のなかでも細胞核の大きさが異なり,また異なる器官で核の大きさがちがうことを確かめた.たくさんの核を大きさにしたがって分類してその体積をしらべてみると“ある基本的な量をもとにしてその整数倍”になっている.この基本量はG. Hertwig (1933)によれば各々の動物の種類によって特異である.核は有糸分裂をなさずに,空間的内容の倍加によって成長するのである.
平均細胞Regelzelle,すなわち固定し包埋してつくった標本でしらべて最も数多くみられる細胞がもつ核の直径と体積は胸腺リンパ球では4.1µ(36ccµ),肝細胞6.75µ(128ccµg),神経細胞16µ(2,145ccµ)である.
核の倍加成長ということはいろいろの哺乳動物でVoss(Z. Zellforsch., 7. Bd.,1928), Clara (Z. mikr.-1nat. Forsch.,13. Bd.,1928; 26. Bd.,1931)およびHertwing, G. (Sitzber. naturforsch. Ges. Rostock, 3. Bd.,1932), Freerksen, E., (Z. Zellforsh.,18. Bd.,1933),また人間では中枢神経系の細胞核についてHertwig, G. (Z. mikr.-anat. Forsch., 51. Bd.,1942)によって証明された.
数:多くの場合1つの細胞は1個の核をもつ.しかしその頻度はいろいろであるがときとして2核をもつ細胞がかなり多くみられるのであって,それは例えば桿細胞や脂肪細胞である.それにくらべるとずっと稀であるが,ほかの種類の細胞たとえば卵や神経細胞にも2核をもつものがある.ある種類の細胞では多数の核をもつこと,すなわち多様性Vielkernigkeitがその特徴なしている.たとえば巨大細胞Riesenzellen(脾臓,骨髄,腫瘍にみられる)がそれである.
核の構造:核は核膜,核材,核小体および核液からなる.ほかの3要素からなる内容物(核形質Karyoplasma)をとりまいて膜があり,その膜も欠けることがあるというわけで,核の構造はかなり細胞のそれと似ている.
核膜Kernmembranは多くの場合はなはだうすいのでその存在を証すことがむつかしい.またこの膜が全く欠けていることも多いが,他の場合たとえば神経細胞では染色などしない生の細胞でも核膜をはっきりと見ることができ,また標本作製によってはっきりとあらわすことができる.
両棲類の卵ではまだ成熟していない卵細胞の核(これを胚小胞Keimbläscheという)を解剖用の針を使って取り出して,その膜を破って核の内容を流しだすことができる.この飼うには多数の小さい孔Porenらしいものがあるといわれる.
核材KerngerustはリニンLininとクロマチンChromatin (nucleïn)とから成っていて,千差万別の形をしてあらわれる.これは生きている細胞のなかにも存在しているのだが,そこでは見ることができないか,あるいはごく不明瞭しか見えない.固定することによって初めて明瞭となる.最もふつうにみられる核材の形は蜂巣状ないし網状の配列である.網の節をなすところにクロマチンの大小いろいろの集まりがあって,これは網の節Netzknotenとよばれ,核小体とは区別するべきものである.他の核ではリニンとクロマチンが1本あるいはそれよりも多数の糸の形で存在する.しばしば数本の糸がうねってたがいにからまって核の縁の近くを通っている.またこの糸から数に多少の差があるが,たくさんの側枝がでて,最大種々の枝でたがいに連なった網状物となっていることがある.
その糸の配列をよく見ると,明らかな極的差異polare Unterschiedeがあらわれていることがある.つまり糸がそれぞれ1本のわなをなしていて,わなの頂(曲がりかどのところ)が核の1極すなわち極野Polfeldにむかっており,わなの両端はその反対側Gegenpolseiteにむかっている.個々のわな状の糸は,その数が一定しているが,多数の側枝によってたがいに結びついている.側枝は二次糸sekundäre Fädenであって,それより太い一次糸primäre Fädenとは区別されるのである(Rabl).
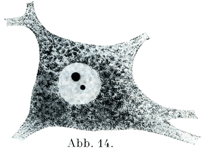
[図14]胞状の核をもつ神経細胞で,大小2個の球状の核小体がみられる.
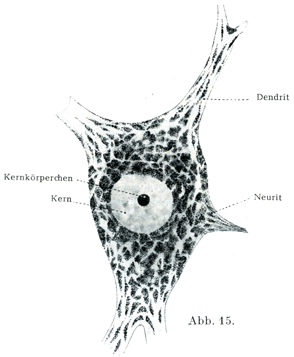
[図15]胞状の核をもつ神経細胞で,核の中央部に1個の球状の核小体がある.
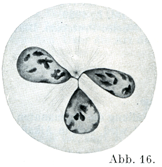
[図16]分葉状の核 サンショウウォの白血球.

[図17]サンショウウォの精巣における休止核のクロマチン糸Chromatinfaden. (W. Flemming)
核小体または仁Kernkorperchen (Nucleioli)は核の内部に1個あるいはそれ以上の数で存在する.これはタンパク質およびごくわずかのリポーゼ核蛋白体RiboseNucleotidenより成っていてふつうの核染色剤によっては染まらない.多くはまん丸い形をしていて,生きている細胞においても明らかにみられる.核剤の成分であるクロマチンといかなる化学的および生理学的な関係を持つかは今日まだよく分かっていない.
Saguchi (Zytolog. Studien 1930,1934)およびBerg (Z. mikr.-anat. Forsch., 28. Bd.,1932)によれば核小体から物質が細胞体に移行するという.
まだ成熟しない卵で胚斑Keimfleckeといわれるものは,若干の理由によって核小体ともよばれているが,ふつうの核染色剤に対するその態度が真の核小体と違っていて,これに染まるのである.真の核小体はそれには染まらない.神経細胞の核小体は特に大きく,染色しない生の標本でもこれを見ることが困難でないのである(図14,15).
核液Kernsaftが特別な性質の液であるかどうか決定するのはむつかしい.核膜に孔があいていないならば,そういう膜は透析力をもつから,核液が特別の性質をもちうるのであるが,核膜のないばあい,あるいは膜が囲んでいても,それに孔のあいているような核では核液は細胞体の原形質とその孔を通して直接につづくわけで,核液と脂肪体の液とが本質的にちがうはずがないのである.
生きている細胞の核内では核小体だけはいつもはっきりと見えるが,その他の諸構造は固定標本で見られるのみで,生の材料ではほとんど見えない.つまり光学的に空つぼoptisch leerであるごとく思える.しかしそれだからといって,2,3の学者が主張するように,核のしめす諸構造がすべて人口産物であるという説にも充分な根拠がないのである.-これについてはWassermann, F., Z. Anat. Entw.,80. Bd.,1926, およびHertwig, G., Handb. mikr. Anat., Bd. I.,1929を比較参照のこと.
化学的組成:核は2つないし4つの核蛋白質より成っている.最も本質的なものが2つあって,それは1. クロマチンChromatin(ヌクレインNuclein)と2. パラヌクレインParanulcein(ピレニンPyrenin).
それになお加えて
3.リニンLinin, 4.アムフイピレニンAmphipyrenin,5.核液Kernsaft.
これらの数種の物質のなかで量においてクロマチンがだんぜん多いのである.
クロマチンは(化学的な)沈殿現象によっていわゆる核染色剤と結びついてつよく染まるものであり,酢酸(1~50%)によって沈殿し,アルカリに溶ける.そしてペプシン塩酸Pepsin-Salzsäureでは消化せず,トリプシンTrypsinでは消化する.
クロマチンという名称は原形質という語と同じように決して科学的に単一な物質に伏せられたものではなくて,1つの形態学的な概念である.それはアルブミンやヌクレインないし核酸Nucleïnsäureがいろいろ異なる量で合してできている.
ヌクレインおよび核酸を含む量が多いか少ないかによって酸性の色素に対する反応が変わるので,A. Fischerはクロマチンの定義を次のごとく述べている.“クロマチンは細胞核の成分をなし,核酸を有してよく染まる物質である.それは核酸を含むことが多ければ多いほど,ますます水溶性の酸性色素に染まりがわるくなる”.クロマチンないしヌクレインの呈色反応はメチル緑の酸性溶液を用いてかなり確実になすことができる.またFeulgenの核染色Nuclealfärbungによっても同じ目的が達せられる(Feulgen, Encykl. mikr. Technik,1927).
パラヌクレインは核小体をなしている物質である.クロマチンとは化学的および呈色的に区別できる.
クロマチンは酢酸(1~50%)によっていっそう明瞭になるが,パラヌクレインよりなる核小体はそのとき膨化する.なおパラヌクレインはトリプシンに溶けがたく,クロマチンを溶かすようなアルカリ性の液に入れても溶けない.また酸性の色素液にいっそうよく染まり,2,3の色素(エオシンEosin,酸フクシンSäurefuchsinをつよくとる.
クロマチンとパラヌクレインとの呈色反応のちがいは次の標語で言い表わすことができる:クロマチンは塩基好性basophil,パラヌクレインは酸好性acidophilである.(この問題ははなはだ多く議論されて,しかも解決していない点が多いのであるが,Fischerの文献をよめばその詳細がわかる).
リニンLinin(Plastin)は細胞核の成分をなす物質で,ヌクレインがその中に宿っているものでる.ふつうの核染色の色素には染まらない.
アムフイビレニンAmphipyreninによって核膜ができている.
核液Kernsaftは多かれ少なかれ核材の網のあいだをみたして存在している.蛋白質を含み,これは固定にあって顆粒の形をなしてあらわれる.
核のない基本有機体kernlose Elementarorganismenが存在するかどうかの問題は,研究方法が精微なものとなるにつれて,ますます否定的な答えの方が有力になってきた.多くの下等の植物や原生動物で以前には核が証明されなかったもので,後にそれが見いだされてきた.微小生物体Mikroorganismenにおいてさえも,その一部では核そのものが証明され,他の一部では少なくとも化学的および呈色的に核の顆粒と同じ反応を示すものが内部にみつかり,その疎とをとりまいている物質が原形質の特色を持つことがわかった.かくして最初はBütschiliがOscillaria, Bacterium lineolaなどで述べたことであるが,KnöllとZapf(Zbl. Bakt.1951)によって細菌もまた細胞核をもつということは確実にされた.
Calkins (Ann. N.-york Akad., XI.,1898)によれば原生動物で最も簡単な形の核は分散型verteilter Kernであって,細胞体内に散らばった顆粒からできている.それより高等なのが中間型intermediärer Kernで,クロマチンの顆粒が1個所に集まっている.核膜はあったりなかったりする.後生動物がもつような核,すなわちリニンではっきりした足場ができていて,その糸にクロマチンが宿っている核は,原生動物でも時たまみられるのである.
細胞の内容をなす何か1つのものが核の性質Kernnaturをもつかどうかをさだめるには,前にクロマチンないしヌクレインの化学的および呈色的反応で述べたところからわかるように,単一の染色方法を用いただけでは充分でない.メチル緑を酸性液にして用いるのは以前はクロマチンを証明する最良の方法とされたが,これさえもFischerによりその価置が疑われている.
クロマチンという名称が形態学上の概念であって,決して1つの化学的物質を意味しないことを前にもはっきり述べたのであって,それ故に細胞の内容をなす或るものが核の性質をもつかどうかを定めるには,大部分の核に共通な形態学的および化学的の性状を全部あわせて考えて,その上で証拠だてるよりほかないわけである.-Kopsh, Fr., Internat. Monatsscher. Anat. Phys., 21, Bd.,1905.-Pratje, A., Biol. Zentralbe. 40. Bd.,1920; Z. Anat. u. Entw., 62. Bd.,1921.
核と原形質との関係:核は一定の作用物質を分泌するという.これをmorphogene Substanzen(形態原物質の意) (Haberlandt, G., Abh. Akad. Wiss. Berlin,1941)といい,それ自身の細胞や他の数多くの細胞の原形質にはたらいて,一定の形態学的性状が発達することの原因をなすのである.この物質がいかなる種類のものであるかについては確かな結果がまだ何も得られていない.
B. 細胞がしめす生命の特徴
R. Virchowがその著わした細胞病理学Cellular Pathologieの中で云っているように,細胞は“真に生命をもつ単位wahrhaft organische Einheit”であって,細胞から“すべての生命活動alle Tätigket des Lebens”がでてゆくのである.それゆえん,われわれは動物の体にみるあらゆる生命現象を,その体を構成する個々の部分すなわち細胞にふたたびみいだすことを当然期待してよいのである.
われわれは細胞の機能に植物性vegetativと動物性animalishのもを区別する.後者は神経性と運動性であり,前者は物質代謝と成長,細胞の生成と増殖,また細胞の分化ならびに寿命に関するのである.
1. 神経性の機能nervöse Funktionen
生きている原形質ははなはだ感受性に富み(sensibel),化学的および物理的の刺激に対して活発に反応する.それは特に温度Temperaturに対して敏感である.あらゆる生命現象の流れにはある程度の熱が必ず伴っているしかしその温度にも適度Optimumと極大Maximumと極小Minimumの区別がある.寒冷は必ずしも原形質の生命活動をやめるものではない.もっともそれはすべての生命活動を抑えて低下させえるのであるが.
動物体内でも熱によって化学作用の速度が増す.10°高くなるごとに2.5倍ほどもはやくなる.(Peter, Arch. Entw.,1905, およびZ. Morph. Anthrop.,1924. )
2. 運動性の機能motorische Funktionen
原形質はすべて収縮する性質をもっている.もっともこの性質の大きさは細胞の種類によってそれぞれちがう.
いままでに知られている運動の形を(a)原形質運動Protoplasmabewegungと(b)鞭毛および絨毛Geißel-und Flimmerbewegungとに分けることができる.
a)第1の群のなかでまた,細胞体の形の変化が起こる運動現象と,細胞の内容物が動いて細胞の形は変わらないものとが区別される.“原形質流動Protoplasmaströmung”は主として固い壁をもった植物細胞におこるのであって,細胞壁に接している原形質の全部がその壁にそって流動すること(旋回Rotation),と細胞の内部をつらぬいている原形質索にそっていろいろの方向に運動すること(循環Zirkulation)とがある.
最も興味があり,同時に生理学や病理学上の諸現象を説明するのにはなはだ重要な運動の形は“アメーバ様の運動amöboide Bewegung”であって,これは細胞の表面に原形質の小さい突起すなわち偽足Pseudopodienができて,これが大きくなったかと思うと,またふたたびそれがひっこめられる.そのあとで表面の他の場所で同じような現象が新たにはじまる.偽足の形成ははじめにほとんど顆粒のない原形質が外方に向かって伸びることで起こる.しばらくして急に押しだされるように顆粒形質のかなり多量あるいは少量がその偽足の方に動いてゆく.その偽足が次にひっこめられないで,細胞の大部分が突起の伸びたところに移るときは,形の変化から位置の変化Ortsveränderungが生じたわけである.この現象は単細胞のアメーバ類で最もきれいにみられるので,それにちなんでこの運動の全部の形がアメーバ様という名前をえたのである.比較的高度な動物の細胞では白血球が最もよくこのアメーバ様の運動をあらわす(図94).
b)鞭毛あるいは絨毛の運動は細胞の表面にある特殊の器官によって行なわれる.表面にある突起の数は細胞の種類あるいは動物の種類によってちがうが,1つのことも2つのこともあり,また数本のことも,はなはだ多数のこともある.この突起は明るい物質からなっていて,かなりの固さをもち,ある一定の方向に打つ性質をもっている.それによって細胞の体じしんが前に進むことになるのは精子や数多くの下等の動物でみられることであるが,あるいは細胞の位置が固定していて,その範囲に達する物質の集団運動を起こすことになる.その非常によい1例は気管やその枝の内部における粘液とその中に含まれる異物が絨毛の運動によって喉頭の方にむかって運びだされていることである.この運動の形が動物体にとって重要な意味を持つことは上述の点からたやすく理解される.
この原形質運動がどんな原因で,そしてどんな機序で起こるかはいまのところ確実なことがわかっていない.機序の問題では特別な糸状の成分があって,これがその運動を仲介して,大事な役目をしていることは間違いないようである(29頁を参照のこと).なおまた,いろいろな種類の筋細胞では細胞の原形質の大部分が特別な分化をとげて,,そこでは運動能力の一般的な性質がはなはだしく高度にそなわっているのである.思うに原形質流動といい,アメーバ様の運動といい,絨毛や鞭毛の運動および筋の収縮といい,みな一群に属して,たがいに深いつながりをもつ現象である.上に述べたいろいろの方向における原形質の収縮性という性質はすべて特別な構造があって初めて実現するのである.もしも原形質が単に液性の物質であったならば,その収縮する性質は理解できないわけであろう.
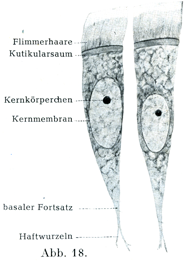
[図18]絨毛をもつ細胞 カエルの口蓋からとりだしたもの.
3. 物質代謝および成長の機能Funktionen des Stoffwechsels und des Wachstums
a)物質代謝Stoffwechsel
物質代謝は物質を摂取し,それを変化させて,また物質を排出することである.それらの物質には気体,液体および固体のものがある.
動物の細胞tierische Zelleに養分として役立つものは,その化学的組成がその細胞を構成している物質に近い一致をもつものでなくてはならない.植物の細胞pflanzliche Zelleでは事情が大いに違っていて,これは合成の仕事を営むようにできている.われわれのみる所では,細胞の栄養という問題で直接にはたらいているのは副形質Paraplasmaであって,これは蛋白質やその他の栄養物質の溶けた液体であり,生命のやどる細胞核や細胞体内の線維のまわりを浸しており,これらに必要なものをあたえ,老廃物をこれらから取り除き,また浸透圧をもひきつづき作っている.
細胞の内部で行なわれる物質の変化は多様のものであって,その一部は生理化学physiologische Chemieの研究すべき領分に属する.他の一部はわれわれが細胞の分化Dfferenzierungをしらべるばあいに,細胞の内部や表面や脂肪の外にみられるものとして,直ちに遭遇するのである.広汎にわたるその変化のなかからここでは次のことだけをあげておこう.その変化に伴ってできたものが細胞をでてゆき,いろいろなぐあいに利用され,あるいは体から外に捨てられる.最後に述べたのは例えば尿素,尿酸,炭酸ガスなどのような多量の本当の分解産物である.しかしまた乳汁のようになお一定の役目をもつ代謝産物もこの分に数えることができる.それ自身の体に特別に利用されるのは腺の分泌であって,例えば肝臓の細胞がつくる胆汁,胃の腺細胞がつくる胃液,唾液腺の分泌物,ならびに皮膚の視線でつくられる皮脂,および種々のホルモンhormoneである.ホルモンは一定の(おそらくはすべての)器官がこしらえる物質であって,それが血管のなかを通って,他の器官を刺激して,その働きを高めたり,あるいは抑えたりする.
電子顕微鏡やその他の方法によって超構造Ultrastrukturがわかってきた.また局所化学Topochemie (Voss, “Mikroskopie” Bd. VII,1952)は構造を保ったままで細胞および組織の化学的組成をしらべるのである.他方,組織化学Histochemieは構造を保つことなく,上と同じ対象を研究するのである.
生きている原形質内で行なわれている化学作用Chemismusは非常にむつかしいことがらである.無機体Anorganismusの化学と有機体Organismusの化学とを区別する主要な標準は何であるかを決定することをC. Wendtが1894年に企てた.(Jen. Z.1894).普通gewöhnlichの化学反応においていつでも必要なもとそして,彼は次のことを数えている.たがいに異なる2種の物質の摂食,溶解する液ないし伝導をつかさどるもの,電気の流れ,体積の変動,熱エネルギーの変化,科学的な集団作用.これらの必須条件のなかの1つだけ,すなわち科学的な集団作用のみが原形質に特有な化学反応には欠けている.それゆえに生活体の反応では化学的平衡状態が起こらない.原子エネルギーが平均して落ちつくということがない,すなわちエントロピーの極大ということが不可能である.原形質に特有な反応は原形質の毛細管のなかで起こるのであって,これが化学者のレトルトのなかで起こる集団反応とちがうのは原子の間ないし原子群(基Radikale)の間の個々の勝負が狭い路でなされるのに,他方は広い戦場で大軍の間で激戦がなされるようなものである.個々の勝負での勝利者が負けた方の原子を従えてゆく,そして限りない複雑さの結合が達せられるのである.つぎに植物原形質Phytoplasmaとして特徴的なものを動物原形質Zooplasmaと比較して,それらの化学作用の主な差異として認められることは,それぞれが違った毛細管引力をもつので科学的取引が2つの相違なる種類となるのである.植物原形質は濃密にし,動物原形質は分裂させる.しかしこれはもちろんごく大体においてのことであって,決してそれのみがおこるのではない.
Hofmeisterはこの問題を別の方向から解こうと試みた.彼は原形質のすでに知られている形態学上の特色から出発するのでなく,その逆に原形質がその化学作用を完全になすことができるためには,それがどんなぐあいにできていいなければならないかという問題を考えてみたのである.肝細胞という具体的な1例にむかって非常にはっきりした,誰でも合点がゆくような考察が加えられた.われわれが今日持っている知識によれば肝細胞のなかで10あまりおそらくもっとそれより多い数の化学現象がごく小さい場所のなかでならんで行なわれている.その場所の広さは止め針の頭の約10万分の1とおもわれるのである.肝細胞は糖からグリコーゲンをつくり,またグリコーゲンから糖をつくる.アミノ酸およびアンモニアから尿素と尿酸をこしらえる.また血液の色素を分解して,それから鉄を分離させてビリルビンをつくる.胆汁酸を生ぜしめて,これをグルココル野タウリンと結合させる.またフェノールと残余硫酸とをあわせてエーテル硫酸をつくるし,なお肝細胞じしんを栄養するために諸現象がある.どうしてこんなにたくさんのいろいろの過程が隣りあって,そんなに小さい場所で起こりえるのであろうか?
それはたがいに反応する物質が溶液の形で遭遇するというかぎりは,原則的としては日常われわれが化学の実験室で行う反応と変わらないである.しかしその反応の多くは触媒的に作用する,ほとんど消費されることのない酵素によって初めて可能となる.これらの触媒物質はコロイドの性質をもっているので,細胞の外に流れ出ることはない.酵素が細胞のもつ化学的用具として本質的なものであることを知るならば,10以上のものそれぞれ違った酵素化学的な過程が同一の細胞のなかで,同時にまたつぎつぎに行なわれうることが理解される.さらに次に述べる2つのことが証明さえているので,その理解はいっそうたやすくなる.1. 同じ1つの酵素が単一の化学物質に働くのみでなく,同じようにできている化学物質の全系列にもはたらく.2. 同一の酵素が条件の変化によって異なるはたらきをする.また個々の反応がたがいにじゃまされずに起こりうるためには空間的に分かれていることが必要であるが,これは酵素がコロイドであり原形質もコロイドであって,酵素が浸透して広がることはほとんどないので,それもはなはだ容易に考えられるのである.それゆえ,細胞の原形質は全体が同じようにできているのではなくて,コロイド性の壁で仕切られた数多くの部屋をもっていて,その壁は部屋の中で起こっている反応に対してかなり抵抗できるものであるにちがいない.この結果は原形質の蜂巣構造の学説とよく合致するのである.Hofmeister, F., Die chemische Organisation der Zelle. Braunschweig. Vieweg & Sohn.
b)成長Wachstum
細胞の物質代謝と直接につづいて細胞の成長という現象がある.もっとも細胞の物質代謝に伴っていつも細胞の増大がおこるとはかぎらない.収入と支出が平衡をたもっていることがある.収入が支出より多ければ細胞の大きさあるいは密度が増すが,その逆だと細胞は小さくなりあるいは密度が減る.細胞の成長は内部から膨れるようなぐあいにおこる(ein intussuszepionelles, inners Wachstum).
4. 細胞の成生と増殖Bildung und Vermehrung der Zellen
古くSchwannが動物の細胞の発生について,Schleidenが植物細胞について最初にのべたのは組織をつくる性質のある液eine gewebebildende Flüssigkeitすなわち細胞形成体Cytoblastemaのなかに自然に生ずるというのであるが,その過程はその後の研究によって誤りであることがわかった.この問題に関係したすべての研究の結果をVirchowが総合して“すべての細胞は細胞より生ずる”Omnis cellula e cellulaという1文にまとめたのである.それと関係してFlemmingの文句“すべての核は核より生ずる”Omnis nucleus e nucleoも直ちにここに付属する.RauberはAltomannやPflügerが“すべての顆粒は顆粒より生ずる”Omne granulum e granuloと主張したのを正しいと考えている.おそらく“すべての中心子は中心子より生ずる”Omne centriolum e centrioloということも正しいであろう.最後にこれから導かれて“すべての生物は生物から生ずる”Omne vivum e vivoとなる.少なくとも今日ではそう云える.
原形質と,それとともに生命がいかにしてはじまったのか,現在のところわれわれは知らないのである.もっともそれについての学説は数多くある.近ごろBalyとButenandtは蛋白質の成分すなわち地球上の生物を構成している物質を人工的につくることに成功した.しかしこれらの物質がいかにして生命をもつに至ったかという問題は今なお未解決である.
すでに存在する細胞がいかようにしてその数を増やすかは,第19世紀の終わりに近い2, 30年の間の研究によって明らかにされた.
細胞の増殖は分裂によっておこるのである.
細胞のすべての成分が分裂する.その減少は中心球および核においてはなはだ目立つのである.それは中心小体と中心球が分裂し,核物質から糸状のものができて,これが分裂する.
この形式の細胞分裂は間接分裂あるいは有糸分裂indirekte od. mitotische Zellteilungとよばれ,それに対して直接分裂あるいは無糸分裂direkte od. amitotische Zellteilungがある.後者は核に複雑な変化が起こらないので,細胞体と細胞核とが簡単にくびれて分裂するのである. まずこれら2種の細胞分裂を形態学的にくわしくしらべてみよう.
A. 間接分裂すなわち有糸分裂
この形式が細胞分裂としてははるかに多く行なわれ,またいっそう重要なものである.それに対して直接分裂は従属的な意味を持つにすぎない.
要領をよくするために,われわれは間接分裂という現象を4つの相次ぐ時期に分ける.その各時期の区切りははっきりしたものでなくて,たがいにいつしか移りゆくのである.1. 前期-中心球と核が分裂のための準備をする時期で,核が動揺をしめし,市場の構造があらわれて,毛毬Spiremがみらえる.2. 中期-核物質が分裂するときで,単星Monasterの時期ともいう.3. 後期-核物質が分配され,原形質の分裂がはじまるとでき,双星Dyaserの時期ともいう.4. 終期-原形質の分裂が終わり,核が休止の状態にもどる.
1. 前期Prophase:休止している核についてまず述べること,このものは核膜,核材,核小体からできている.核の1個(極側Polseite)に軽いへこみがあって,そこの原形質のなかに中心球が1個あるいはすでに2個となった中心子をもって存在している(図19).
中心子の分裂Zentriolenteilungはつぎのように行なわれる単一の球形の中心小体が1本の棒状のものになって,これの中央部が細くなって亜鈴状になり,そしてついに2つの小球に分かれる.そのさい球形の中心球が楕円形になり,これは初めは分裂した2つの中心子をもっているが,ついで中心球もまたつに分かれて,その各々が各1つの中心子をもつものである.
核の染色されやすい物質はまず1本,2本あるいはそれより多数の糸を形成し,これが核膜のなかでうねってもつれたようになっている(毛毬Spirem).毛毬は初めは密であるが,次第に疎になる.そしてこの糸が染色体Chromosomenとよばれる一定した数の断片に分かれる.
染色体の数と形Zahl und Gestalt der Chromosomenは動物の各種ならびに細胞の種類によって違っている.球形,卵円形 輪状,棒状,まっ直ぐもの,曲がったものがある.比較的高等な動物で普通にみられる形は係蹄(わな)である.婦人の髪を止める針Haarnadelに似ている.それゆえ,染色体はまた核係蹄Kernschleifenともよばれる.その数は動物の種によって定まっており,特徴的であるといわれる.しかし,比較的筋人柄の研究によって同一の種類の動物でも若干の変動があることがわかってきた(Karplus, Z. Zellforsch.,10. Bd.,1929).人間の細胞はおそらく48の染色体をもつのである.Flemming, duesberg, Grosser, Schachov, Tage Kempは24であると云った.マウス,サンショウォ,マスは24である.ニワトリは(Saguchiによれば)30,軟骨魚類36,ウマに寄生する蛔虫Ascaris megalocephala univalensは2,同じくAscaris megalocaphala bivalensは4である.Duesberg, J., Anat. Anz. 28. Bd.,1906.-Grosser, O., Verh. anat. Ges., Marburg ,1921; および“Der Deutsche Arzt”1938.
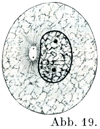
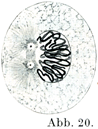






[図19~26]間接分裂をしめす模式図 中心子のまわりの明るい部分が中心球である.前期は図19, 20, 21, 中期は図22, 後期は図23, 24, 終期は図25, 26.
染色体の微細構造は細菌の10年間にかなり精しく研究された.染色体は決して無構造のものではなくて,クロモネマChromonemaというラセン形にうねった糸状物よりなりこれにクロモメーレンChromomerenという無数の小体が付いている.後者は遺伝原基をもつものであるといわれる.
核係蹄ができるあいだに核膜また消える,そして核液が細胞体の液と混じり合う.しかしそれでも核のあった場所と原形質との境は保たれているのである.それと同時に2つの中心球はたがいにいっそう遠ざかり,前期の終わりになると,核の周縁でたがいにそう反するところに位置するようになる.その2つの中心球のあいだは紡錘状あるいは樽形のものがあって,これは軽く弓形に曲がって走る染色されがたい細い糸からできていて,不染色性の中心紡錘Zentralspindelとよばれる.他方また,各々中心球のまわりにあらゆる方向に広がる放線状の細い糸がみられて,これが極放線Polstrahlungあるいは星Asterである.極放線の大部分は細胞の周縁のところで,原形質の網状をなす構造のなかに消えるか,またはそこにいくらか存在している細胞膜に直接に付着している.他の部分は染色体に結合していて,これが牽引糸Zugfasernとよばれる.これはそれはこの糸が染色体を2つの娘細胞に分配することをつかさどると考えるためであるが,またこれが中心紡錘のまわりをとりまいて存在するので,外套糸Mantelfasernという名前もある.中心紡錘の線維は2つの中心球が近づかぬようにささえるはたらきをもつと考えられているので,この理由から支持糸Stemmfasernと称せられるのである.
2. 中期Metaphase(図22):中心紡錘の形成は最高潮に達していて,その長軸は細胞じしんの長軸と一致している.染色体はそのあいだにいっそう短く太くなり,係蹄の曲がりかどを中心紡錘にむけ,係蹄の両脚を細胞の周縁の方に向けるように位置する.この時期の細胞を1極からみると,染色体の集まりが1つの星の形をしている.これを母星Muttersternあるいは単星Monnasterの時期という.同時に染色体は中心紡錘に垂直の一面上にある.この面は赤道面Äquatorialebeneとよばれる.赤道面はあとに細胞体の分裂が起こる場所にあたっている.
ここの染色体(母係蹄Mutterschleifen)がいまや縦裂して全く等しい2つの部分すなわち娘係蹄Tochteschleifenに分かれていることが明瞭となる.この縦裂はすでに前期の終わりにはじまっていたのであるが,この時期になると中等度の拡大でもはっきりと認められるのである.
係蹄が分裂する現象をくわしくみると,母係蹄をなしている多数のクロマチン小球がおのおの伸びて,ついで全く等しい2つの部分にわかれて,そのおのおのがそれぞれ娘係蹄をなすのである.
3. 後期Anaphase(図23, 24):娘係蹄はそれに付着している,そして収縮する性質のある牽引糸(すなわち外套糸)によって一方の中心球へ引き寄せられる.そのときに中心紡錘の線維は収縮する性質を欠き,支持糸として2つの中心球がたがいに近づかぬように支えている.れにまた,中心球のまわりから発して細胞の表面に付着している放線もその支持にあずかるのである.これが娘星Tochtersterneあるいは双星Dyasterの時期である.両極に向かってはなれつつある2群の染色体の集まりのあいだは結合糸Verbindungsfadenという細い糸でたがいに結びつけられているようである.結合糸はたぶん中心紡錘の線維にほかならないであろう.
4. 後期Telophase(図25, 26):染色体は中心球のすぐそばまで達して,短く太くなり,たがいにいっそう密に寄り合う.そしてたがいに結合して,いままでその表が平滑であったのが,小さい突起を出すことによって,ギザギザとなり,でこぼこをもってくる.そして結局,核膜によって包まれる.次第に変わってゆきふたたび小胞状の核ができるのである.その変化は核が前期になしたものでを,今やその逆の順序で行うのである.そのあいだに中心球のまわりの放線は失われてゆき,それが全く消えてなくなることもある.しかし中心子はしばしばこの時すでに2分している.
細胞体の分裂はこの時に初めてはじまる.もっともときとしてはそれがもっと早く娘星のときにはじまることもある.赤道のところの1個所に溝ができてはじめて,これがついで赤道をまわる輪の形になり,鱗状のくびれができあがる.このくびれが深くなって,ついに細胞体の分離がおこるのである.原形質の分裂が起こらないと,2核をもつ細胞ができる.
いろいろな長さの休止期をおいて,しばしばその休止期ははなはだ短いこともあるが,核は新たに動的な時期,すなわちつぎの分裂にはいってゆく.そのさい注意に値するのは次いでおこる分裂の軸がそのすぐ前におこった分裂の軸に対して直角をなしていることがしばしばであり,また相ついでおこる分裂の軸が空間の3次元に相当するというぐあいである.しかしこれにも類型的な差異typische Verschiedenheitenがたくさんにある.
G. Hertwig (Sitzber. naturf. Ges. Rostock,1930)は染色体が律動的な成長をなして,その量が倍加すると考えた.1価の染色体から2価のものができ,また後者が倍加をくりかえして4価およびそれ以上の価の染色体ができる.そういう倍加成長をした各時期の終わりには染色体は分裂する能力を持ち,それを含む核は分裂の用意ができている.そのときに本当に分裂がおこるかどうかは他の因子によって決まるのである.温熱や冷却やコルヒチンColchicinのような化学物質によって染色体の数が何倍にも増したりする(倍数性Polyploidie) (Häggquist, G.,1948, Häggquist, G. und Bane, A., Nature, Vol.165,1950, Melander, Hereditas, Vol. 36,1950).
有糸分裂のあいだは(Meves, Festschrift Kupffer,1899による,また特にPeter, K., “Protoplasma”,10. Bd.,1930)その細胞のもつ多種多様の生活現象が中絶して,ただその細胞が分裂の時期を生きながらえることができるだけの生活機能しかなされていない.これに反して無糸分裂は細胞の活動を害することがない.
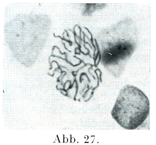

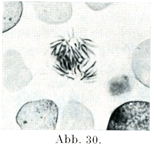

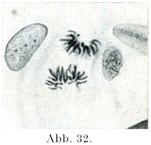

[図27~33]サンショウウォSalamandraの表皮細胞における核分裂像および休止核 図27は前期,図28, 29は中期,図30~32は後期で,双星の時期,図33は終期.
有糸分裂に要する時間Dauerは動物の種類によって異なる.それは温血動物では冷血動物におけるより短いのであって,人間ではおそらく30分間であり,サンショウウォでは温度によってちがうが2~5時間(Flemming)という.Peterはこの2~5というのが少し短く見積もりすきていると主張した.ダツ(硬骨魚)Belone acusやネメクジウォ(頭索類)Amphioxusの卵の分割では(18~19℃で)約1時間,硬骨魚のGobius(ハゼの類)やCrenilabrusの卵の分割では(15~19℃で)30~40分である(Fr. Kopsch).有糸分裂の日時的変動Tagesschwankungenがオタマジャクシの表皮の細胞において証せられている(Möllenberg,1948).
温熱は細胞分裂の進行を早くする.両棲類では温度が10℃あがるとその速さが2倍になる(Peter, K., Z. Morph. anthrop. 24. Bd.,1924).しかしこれは一定の好適な温度内のことであって,温度がそれより低いときは遅延因子,それより高いときは加速因子を加えなければならない.いろいろな侵害が有糸分裂の新興に影響をあたえ,あるいはそれを抑制する.例えば比較的高い温度(42℃~45℃) (Wassermann, Verh. anat. Ges.1921), Kokott, Z. Zellforsch.,11. Bd.,1930),冷汗,麻薬薬(Politzer, Z. Zellforsch.,13. Bd.,1931),酸をはたらかせること,気圧を何倍かにすること,レントゲン線(Politzer, Strahlentherapie, 42. Bd.,1931), 細胞の死が,いま云ったような侵害として数えられるのである.細胞分裂は創傷ホルモンWundhormoneあるいはNekrohormoneによってもひきおこされうる(Haberlandt, Biol., Zentralbl., 42. Bd.,1922).これは外傷により破壊された細胞あるいは単に傷害されたか,ないし死滅した細胞からでてくる物質である.また細胞分裂はミトゲン線“mitogenetische Strahlen”によってもひきおおこされる(Gurwitsch, Das Problem der Zellteilung. . . Berlin,1926),これは“Organismentrahlung”(Stempell) (有機体放射線の意)というのがいっそう適切である.これは紫外線であって,その波長はこれを発するもの(放射源StrahlungsquelleあるいはInduktor)の如何によって違っている.
分裂を起こす因が核にあるのか,あるいは原形質か,あるいは中心球にあるのかという問題で学者たちの意見が一致していない.中心球が指導的な役目をしているごとく一応はみえる.しかしこの3つのものの交互作用の上に分裂をおこす因があるとの考えもいま述べたのと劣らず脳裡におく必要があるのであろう.
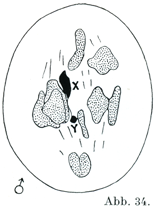
[図34]ヒトの精母細胞SpermiocyteにおけるX-およびY-染色体(Painter, J. exper. Zool., 37. Bd.,1923による.)
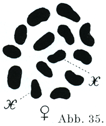
[図35]半翅類カメムシLygaeus turcicusの卵祖細胞Oogonieにおける1組の核係蹄.

[図36]Lygaeus turcicusの精祖細胞Spermiogonieにおける1組の核係蹄

[図37]Lygaeus turcicusの精母細胞Spermiocyteの第2分裂におけるX-およびY-染色体.(図35~37はEd. B. Wilson, J. exper. Zool., 3. Bd.,1906による.)
常染色体Autochromosomenと異染色体Heterochromosomen
多数の動物および植物において(とくに昆虫で明瞭であるが)1個の細胞のなかにある染色体が同じ種類ではないのであって,大きさ,形,そして(たぶん)意義が違っている.いっしょにして染色体の組Chromosomen-Garnitur, ドイツ語でKernschleifensatz(1組の核係蹄の意)とよばれる.
1個の細胞のなかにある大きさや形のちがう染色体をよく調べてみると,2つずつ同種であって,有糸分裂のときにこの2つがしばしは対をなしていっしょに存在している.これが相同の染色体homologe Chromosomenとよばれる.ところが動物によると,こういう対をなす常染色体AutochromosomenあるいはEuchromosomen(正染色体の意)のほかになお1個の特別な染色体があって,これは対をなす相手をもっていないか(そのばあいX染色体X-Chromosomという),あるいは対をなすがその相手と大きさも形もちがうのである(この相手はY-染色体Y-Chromosomとよばれる).このXとYを合わせて常染色体に対して異染色体HeterochromosomenあるいはIdiochromosomen(特異染色体の意)という.これが性の分化に関係を持つといわれ,したがって性染色体Geschlechtschromosomenともよばれる.しかしこれが性を決定するということがまだ確定されているわけではない.
Karplus (Z. Zellforsh.,10. Bd.,1929)は人間において1組の核係蹄がいつもその染色体の1つ一つが形と大きさが定まっていることを証し得なかった.また人間では異染色体を見つけることができなかった.(人間における異染色体の存在は今日では確立している(小川鼎三).)
B. 直接分裂あるいは無糸分裂direkte oder amitotische Teilung
Flemmingの定義によれば無糸分裂というのは細胞分裂および核分裂の一型であって,これには紡錘の形成がなく,染色体が規則正しくつくられることもなく,また染色体が定まった形および順序に位置を変えることもないのである.
1個の細胞とその核が亜鈴状にくびれて,そのあいだを連ねる部分が細長く伸びて,そこがついに切れる.または1個の細胞が2つの核をもつようになって,そのうちの1核が原形質の一部とともに芽が出るように残りの部分から離れる.白血球ではこのような無糸分裂がみられる(図38).もっとも白血球では有糸分裂もおこるのである.
有糸型の核分裂はとくに受精卵から胎児ができてくる途中では圧倒的に多くみられるが,個体の生存する全期間を通じてもやはりこれがはなはだ多く見られるのである.無糸分裂はそれに比べるとずっと稀である.Krompecher (Z. Anat. Entw.,107. Bd.,1937)およびPfuhl(同上誌,109. Bd.,1938)は,有糸分裂はまだ1つの方向にある程度以上に分化をとげていない細胞におこるもので,それに反して無糸分裂は1つの方向に分化をとげた細胞の核の最終的な分裂であると論じた.同様な意見はすでにFlemmingおよびZiegler (Anat. Anz.,12. Bd.,1896)が述べたところである.
Hoepkeによればコウモリやハリネズミの脾臓の細胞では有糸分裂が圧倒的に多くみられる(Verh. Ges. Naturw. Marburg,1922, およびStöhr. Z. mikr.-anat. Forsch., 36. Bd.,1934を参照のこと.-Peter (Z. Anat. Entw., 75. Bd.,1925)によると,無糸分裂は“細胞に分裂の必要が迫って,しかもその細胞の特殊な活動を中絶できない”立場にあるときにおこるのである.無糸分裂は細胞の活動をじゃますることがないのである.(Peter, Z. Zellforsch. 30. Bd.,1940).
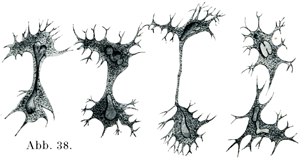
[図38]カエルの白血球における直接分裂.(Arnoldによる.)
5. 細胞の寿命Lebensdauer der Zellen
受精した卵細胞から出発して,ひきつづき分裂がおこなわれて,はなはだ数多くの細胞ができて,これが組織や器官にまとまって生物体をなすのである.そのほんの一部にすぎない少数のものが生命の繁殖をその任務とする胚細胞Keimzellenとなる.そしてそれを除いた他のすべての細胞は遠かれ近かれ死滅するのである.しかもすでに胎児の時代にもいろいろな具合で,細胞の真の破滅がおこるのであって,これは全く生理的な現象である.例えば上皮細胞が剥げてゆくこと,,腺細胞が溶けて死ぬこと,胎児や臍帯といっしょに胎児被膜が外にだされること,骨化のときに軟骨組織が壊されること,骨の成長に伴って骨の吸収がおこるときに骨組織がこわされること,軟骨発生にあたって骨の吸収がおこるときに骨組織がこわされることなどである.またそれほどめだつ程度でなくても生涯の全体を通じてみると,たとえ毛の脱けて落ちることや月経のときの出血のような著しい高度の損失を除外視しても,古くなった細胞が滅びてゆくという現象は多くの種類の組織でおこなっている.そのさい病的な破滅もしくは消滅の明瞭なしるしはないのである.
それゆえ,非常に数多くの細胞がそれが属する個体よりも早く死ぬのである.その属する個体と同じだけ長く生きる,すなわち個体発生の初めからその死滅まで生きるのはどの種類の細胞であるかという問題は当分まだ確実には答えられないが,おそらく神経組織がそれであろう.
生物体を成している細胞の大部分はその個体が死んだ後もなおしばらくは生命を続ける.それも生物の種類と細胞の種類によって様子がかなり違う.温血動物の細胞で,物質代謝および熱の影響からみはなされた細胞は最も早く死んでしまう.
ここの細胞の老衰と死滅は一部は原形質に,一部は核にあらわれるのである.死にむかいつつある細胞でも核がなお或る段階の運動をしめす.目前にせまっている破滅がいわば刺激としてはたらくのである.死滅に近づいた卵胞上皮細胞では核のクロマチンが密集して球状のものになる.ついで核の細胞体に対する境がくずれて,後者のなかにクロマチンの塊が散らばってみられる.原形質のなかにおそらく脂肪より成ると思われる多数の微細な小滴があらわれる.最後に細胞体がこわれて,核の破片が卵胞液のなかに溶ける.この現象をFlemmingは染色質溶解Chromatolyseとよんだ.細胞の死滅するいま1つ別の行き方は原形質の水腫様変化Hydropisierungであって,その例は成長し続ける骨の骨化帯のところの軟骨細胞である.また別の行き方が角化Verhornung, Keratinisierungであって,表皮が小鱗をなして剥げておちるのがそれである.なお脂腺の細胞のように脂肪化することFettmetamorphoseもあり,あるいはその他でも腺の分泌物をつくるために細胞が消耗して死滅することがある.
正に死につつある生のままの細胞の内部でどんな化学変化が起きるかをしらべてみると,生命の停止と同時に分解Zersetzungenがはじまる.それは腐敗とは別であり,腐敗を起こす微生物の存在を要しないのである.グリコーゲンやその他の含水炭素を変化させる酵素があらわれる.ついでクロマチン(ヌクレイン)の分解がはじまる.しばらくすると,死んだ細胞の内部あるいはその近くに結晶があらわれるが,これは主にコレステリンと脂肪酸からなっている.ときとしては分解現象が深く進んだ結果としてロイチンLeucinやチロジンTyrosinがでてくる.(A. Kossel. )
何世紀も前の文学者が老衰に関してつぎのような適切な表現をしている:Hebescunt sensus, torpent membra, visus, auditus, incessus, dentes. Sicmagna pars mortis jam praeteriit; quod reliquum est mors tenet. (鈍くなる感覚,麻痺する四肢,目も耳も障害をうけて,歯も同様である.死のそんなに大きい部分がもうそばを通っている.すなわち残るのは死の到来のみだ.)
Bichat, X., Über Leben und Tod.1802.-Goette, Al., Über den Ursprung des Tode S.1883.-Weismann, A., Uber die Daucer des Lebens; Über Leben und Tod.1882および1884.-Merkel. Fr., Über die Gewebe beim Altern. X. Internat. Kongreß. 2. Bd.-Mühlmann, M., Über die Urasche des Alter S.1900; およびErgebn. Anat. Entw., 28. Bd.,1929.-Westergaard. Die Lehre von der Mortalität und Morbidität. 2. Aufl. Jena,1901.-Doms, Herbert, Über Altern, Tod und Verjüngunng. Ergebn. Anat. u. Entw., 23. Bd.,1921.-Korschelt, E., Lebensdauer, Altern und Tod. 3. Aufl. Jena,1924-Bürger, M., Altern und Krankheit. G. Thieme, Leipzing 1974.
6. 細胞の再生Regeneration der Zellen
生理的あるいは病的に生じた物質欠損を補充することによってどんな程度にでも元の状態に復そうとする力を再生Regeneration,またはReparation(修理の意)というのである.
細胞じしんの再生zellulare Regenerationということああるだろうか? つまりここの細胞がその身に物質欠損を受けたときに修理できるかどうか,わわれの経験はそれについて肯定の答えをあたえる.次の観察をあげておこう.
滴虫類Infusiorienの体を人工的に切って実験すると核をもたない部分は生き続ける力をもたないようである.それゆえ,M. Nussbaum (Arch. mikr. Anat., 26. Bd.,1886)はつぎのごとく述べた.1核と原形質はそれらがいっしょになっていてのみ生きてゆくことができる.それぞれ孤立すれば,多少の時間の差はあるが死んでしまう.2. 核は細胞がその形をつくってゆくエネルギーformgestaltende Energieを保つために欠くこのできないものである.
卵細胞から核を取り除いたものを人工的に受精させることができる(すなわち1つの核をもつわけである),そうすると分裂が起きて胚が発生する(Driesch, Boveri, Delage). Schmitzはミドリゲ類Siphonocladiaceen(緑藻類に属す)の細胞をばらばらにこわしてその個々の断片についてしらべると,すくなくとも1個の核をもつ断片だけがその後に生きつづけて,独立した新しい細胞にあることができるのを知った.Strasburgerたちも同じような他の例を述べている.やはり緑藻類の一族であるイワヅタCaulerpaでは驚くべき再生能力が証せられている.この植物は沢山の核をもつ単一の細胞である.すなわち多核の巨大細胞ということができるし,またいくつかの根,1本の匍う根茎Rhinzom,茎や葉までもった1個の合胞体といえるのである.ところがこの植物に生殖器は今日まで何もみいだされいない.その繁殖は自然に,あるいは人工的に断ち切られた部分が再生することのみによっておこるのである.
比較的高等な動物や人間においても個々別々の細胞Einzelzellenの再生量句はかなり多く知られている.もっともよい例は神経細胞である.生きている体で神経細胞の突起が切断さえると,その中心端すなわちその突起をだしている神経細胞とづづいている切り口の方が次第にのびてくる.そしてかなりに行き届いた修理が起きるのである.他の種類の再生としては細胞の全体,細胞団の全部,はなはだしいのはさらに異なる種類の組織の集まりが死滅したために生じた物質欠損を補う場合である.
このばあい傷をうけた細胞じしんがその物質欠損を治すことはできないので,その固体が残されてもっている傷ついていない健康な細胞,しかもその傷の誓うに或る細胞が修理の役目をする.例えば体の表面から表皮の細胞がたえず剥げてゆくが,この剥げる細胞じしんが再生することはできないので,その体に残っている細胞が分裂によって新しい細胞をつくり,これが物質欠損を補充するのである.このばあいの再生にも正常のと病的のとあって,後者は人工的にできた大小いろいろの物質欠損のさいにおこるのである.
それぞれの場合に再生能力の大きさがどれくらいかという問題がおこる.1本の指,1本の腕,胴の一部,1個の目が除かれたときはどうであろうか?動物によるといまいったすべての欠損を再生によって治しうる.分割をなしつつある卵から1つの分割球を取り除くことができるが,その残りから完全な形の肺が生じる得るのである.それでもわかるように,再生の範囲ははなはだ広く,かつ重大である.
Barfurth, D., Regeneration und Involution. Ergeb. Anat.1903およびそれにつづく年次の同誌.-同じ人,Regeneration und Transplantation in der Medizin.1910.-Korschelt, Regeneration und Transplantation. Jena.1907.-Strasser. H., Regeneration und Entwicklung. Jena ,1889.
1-04

最終更新日 13/02/03