Rauber-Kopsch解剖学
歴史的な偉大な解剖学書
Rauber Kopsch Band2. 30 


S. 287
V.神 経 系Systema nervorum, Nervensystem
A.神経学総論
1.序論
すでに前に載せた体の横断模型図において,神経系および感覚器系は2つの環としてあらわされている,すなわち神経系の環(図1, 2)と表皮性の環(1)とである.
神経系がその作用を営みうるために,この系統は身体の他の部分と密接な関係におかれ,かつ身体のなかにはなはだ広くひろがっている.
この点では脈管系と似ているし,そのうえ多くの場所では脈管系と共同の路を通っている.そして脈管系にも中枢の部分と末梢の部分とが区別されるように,神経系でもまた同様の区別がある.脈管系じしんが神経を豊富に受けている.
中枢の部分,すなわち中枢神経系Systema nervorum centrale, Zentralnervensorstemは脳および脊髄よりなる.末梢の部分,すなわち末梢神経系Systema nervorum periphericum, peripherisches Nerwensyrstemには末梢神経およびこれとつづいている末梢の神経節Ganglien, Nervenknotenがある.神経節にはさらに2種類がある,すなわち脳脊髄神経節cerebrospinale Ganglienと交感神経節sympathische Ganglionである.交感神経節はこれらの神経節相互のあいだを結びつけたり,また脳脊髄神経系とこれらとの間をむすびつけている多数の細い神経,およびこれらの神経節から出る細い枝とともに交感神経系sormpathisches Nervensystemあるいは植物神経系vegetatives Nervensystemまたは自律神経系autonomes Nervensystemというある程度独立した系統,いわゆる交感神経Sympathicusをなしていて,これは内臓,腺,平滑筋,血管およびおそらくは横紋筋にも分布している.
2.神経分節neurales Segment
神経分節には次に述べるような諸型がある:すなわち中枢器官Zentralorgan(図355)から各側1本の腹方の運動性の神経根motorische Nervenwurzelすなわち前根Radix ventralisが出て,これは側方に向かっている.この神経根に対して背方の,本質的には知覚性である神経根sensible Nerwenwurzelすなわち後根Radix dorsalisがやはり1本あるが,この後根には脊髄神経節Ganglion spinaleという1つの膨れた神経節がある.前根はこの神経節にただ接しているだけである.この神経節の末梢がわで,これらの両根が合して脊髄神経Nervus spinalis, Spinalnervの短い共通幹となり,これがさらに4本の枝に分れている:すなわち
1. 後枝Ramus dorsalis;これは後方に向かってすすみ,内側の枝と外側の枝とに分れて背部の筋肉および皮膚に分布する.
2. 前枝Ramus ventralis;これは体壁のなかを腹方に走り,次の2本の枝を出す.a)外側皮枝Ramus cutaneus lateralis,これはさらに1本ずつの後枝と前枝とに分れて体の外側部の大きな帯状の皮膚区域に分布している,b)腹方の前皮枝Ramus cutaneus ventralisは正中線に近接する前面の皮膚領域を支配している.そのうえに前枝は特に四肢の筋肉をも含めた腹方の筋肉全体を支配するのである.
3. 硬膜枝Ramus meningicus;これは細い枝であって,次に述べる交通枝Ramus communicansから細い小枝を受けとった後に,椎間孔を通って脊柱管のなかに入り,脊髄とその被膜ならびに脊柱管の壁およびこの管の中にあるその他の内容物に分布している.
4. 交通枝Ramus communicans;この枝は特に内臓と血管とを支配し,かつ交感神経の幹とつながっているが,また交感神経からの線維束を脳脊髄神経系のなかに導いてもいる.
神経分節が上に述べた形をなしているのは身体のなかで胸部だけである.腰部ではすでに下肢が存在するためにある程度の変化がおきている.頚部および特に頭部ではこの変化がさらに高度である.
3.神経系の構成要素およびそれらの連絡関係
神経系は主として神経組織よりなっている,すなねちニューロン(神経元,神経単位)Neuronとグリア(神経膠)Neurogliaとからなるが,なおその構成には結合組織と筋組織とがあずかっていて,この両組織は血管およびリンパ管として多量に存在するのである.
これらの構成要素のすべてについてはすでに第I巻で述べてある.
ここでは器官としての神経系の構造Organstruktur des Nervensystemsについての重要な点を明かにするために,個々の要素の連絡とその関係についてなお少しばかり書き加えることにする.
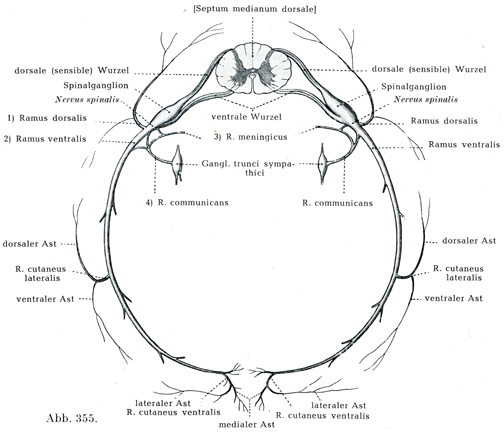
[図355]ヒトの中枢および末梢神経の横断
a)神経細胞Nervenzellen
ニューロンをその突起の数によって単極unipolar,双極bipolar,および多極multipolarのものに区別する.もう1つ別の分類では神経突起すなわち軸索突起Neuritenが長いか短いかが,区別の目じるしとなっている
:すなわち
α. 長い神経突起(軸索突起)をもつ細胞;その神経突起がしばしばその細胞体のそばでいくつかの細い側枝Kollateralenをだしたのちに1本の有髄神経線維の軸索になる.長い突起をもつ細胞といっても,その神経突起(軸索突起)の長さがはなはだまちまちである.人ではその最も長いものは1メートルをこえることがある.
β. 短い神経突起(軸索突起)をもつ細胞;このものではその神経突起が細胞体の近くでつぎつぎと分岐して,間もなく神経の枝の密な集まりとなって終わっている(第I巻,図119).
b)神経線維Nervenfasern
中枢神経系の神経線維は一部は有髄markhaltig,一部は無髄marklosである.これらの神経線維は,シュワン鞘Schwannsche Scheiaeをもたずにグリアに取りまかれているという点で,総論の中に記した神経線維の構造と違っている.
c)神経線維フエルトNervenfaserfilzおよび神経網Nervennetz(基礎網Grundnetz) (図356, 357).
われわれはすでに組織学において,灰白質の内部にはニューロンの突起,しかもその樹状突起ならびに神経突起(軸索突起)とその側枝がつぎつぎに分れて枝を出して終末分枝Telodendren, Endbdiumchenを成していることを知ったのである.この終末分梗の最も細い枝は結局ただ1本の神経原線維といくらかの原線維周囲物質Perifibrillarsubstanzだけからできている(Bethe).
以前には一般に次のように信ぜられていた.隣り合せている分枝構造は直接にたがいに続いており,神経の中枢を貫いて広くひろがり,到るところにはいりこんでいる神経網Nervennetzが存在するということであった.
次いでゴルジの染色法Golgische Methodeによってえられた所見およびこの所見を基礎にして樹てられた神経元説Neuronenlehreの影響のもとに,ニューロンが直接に続いているということが否定されるようになった.それより古い時代の学者たちが神経の網と考えたものは数多くのニューロンの分枝が噛み合うことによって生じた緻密な神経線維フェルトNervenfaserfilzにすぎないと一般に理解されたのである.ニューロンの間の互いの結合(シナプシスSynapsのは,生理学的観察ではその結合が必ず存在するようであるが,組織学では両方のニューロンが相接したがいに触れていることによってのみ起る.といわれ,あるいはまた両者が2次的に合一することによって起るというのである(Golgi, Kölliker).
さらにその後になって,神経原線維を染め出したものを基にして,格子状をなす真の神経網の存在がふたたび記載されるにいたった.これはHeldによって汎在基礎網“allgemeines Grundnetz”として人の小脳の灰白質および白質のなかで記載されたのである.
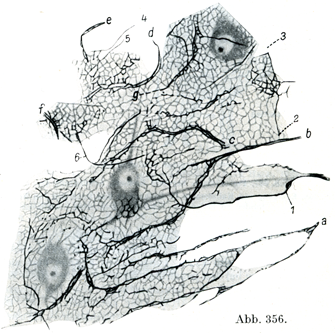
[図356]汎在基礎網 ヒトの小脳皮質.a~g樹状突起,1~6 神経突起(軸索突起),その神経原線維は基礎網のなかにある.(Held,1929)
その構成には神経突起(軸索突起)および樹状突起の最も細い末端部ならびにグリア網胞のそれが関与している(図356).それゆえ脳皮質は神経合胞体Neurencytium(Held)である.同様な所見が人の大脳皮質についてもK. Bauer(1940)によって得られている. Bauer(1943)はこの構造を綜合器官Integrationsorganと名づけ,かつ人の大脳皮質では動物の脳におけるよりもこれがはるかによく発達していることを指摘している.
Bauer, K, F., Organisation des Nervengewebes und Neurencytiumtheorie. München 1952.
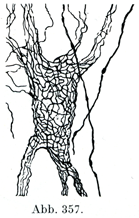
[図357]細胞周囲の神経終末網 若いイヌの神経細胞(Veratti,1900).
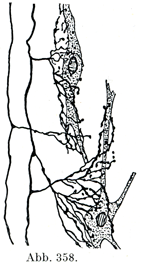
[図358]ニューロンの細胞体および樹状突起に側枝がきて終る様子 後索の線維であって,その側枝が後柱膠様質の細胞に達しているのである(Betheの原著, Cajalより引用).
これらの事実はニューロン説に賛否両方の多数り議論を生むこととなった(Nissl 1903, Bethe 1904, Schiefferdecker 1904, Strasser 1907, Held, H., Fortschr. naturw. Forsch. Berlin 1929).ここではその詳細を述べることができないので,この問題に興味をもつ人には上に述べた諸学者の書いたものを読むことをすすめる.--W. Kirscheは(Psychiatr. Neur., med. Psych., 6. Jhrg. において)ニューロン説を固持している.シナプシスは時により変りうる生きた構造であるという.--Harrison(Arch. exper. Zellforsch., 6. Bd.,1928)によれば培養した神経組織での観察はニューロン説に味方するものであるという.--Bauer, K., F., A. Psych. u. Z. Neur.1951.
学生を教えるに当たって,神経系の概観を得させるためには,個々の神経網胞とこれらから出るすべての突起を合せたものに対してわれわれは総括的な名前をもつのが都合がよいのであって,それにはニューロンという名前がやはりいつでも適当である.たとえすべてのニューロンが中枢神経系の汎在基礎網の中で,あるいはまたその他の場所でたがいに続いているとしてもである.
それゆえこれより以下ではニューロンという語は「1つの神経細胞とそのすべての突起およびその終末までもふくめたもの」という意味をもつことにする.
d)細胞周囲の神経終末網Perizellulares nervöses Endnetz(Held)
いろいろなニューロンの間の結合は汎在基礎網のなかだけで起るのでなく,1つあるいはそれ以上のニューロンの細大種々の枝が別のニューロンの細胞体や樹状突起に接していることが広くみられるのである.細かい線維からなる網目構造,すなわち線維籠Faserkorbが細胞体およびその突起の周囲にあって,この網には外からかなり太い枝(他のニューロンに属する側枝や樹状突起)が入ってきている(図358).
この網目の比較的あらいものからいっそう細い線維が出て,これが神経細胞の細胞体および樹状突起の表面に目の細かい網であるゴルジ外網äußeres Golginetz(図357)を作っている.この網はごく少量の原線維周囲物質に包まれた神経原線維より成り,細胞体および樹状突起に外からぴったりと接している(Bethe). Heldによれば(図359)神経細胞にやってくる多数のごく細い神経枝の終末が肥厚して神経終末足Nervenendfüßeをなし,これが突起を出してたがいにつながっている.
こうして細胞周囲の神経終末網Perizellulares nervöses Endnetz(図359)が生ずる.この網の結び目のところが神経終末足であって,ここには格子状の神経原線維網があり,原線維を神経細胞の内部に送りこんでいる.その包まれている神経細胞から原線維が細胞周囲の神経終末網の中に出ていることも当然考えられるのである.この網は外方へはいわゆる汎在基礎網“allgemeines Grundnetz”と続いている.従って細胞周囲の神経終末網はたんに基礎網の一部であるに過ぎない.
それゆえHeld(1927)は灰白質の中で神経原線維の格子を次の3つに区別しているのである:すなわち1. 神経細胞の内部にあるもの,2. 細胞周囲の神経終末網,3. 汎在基礎網.
したがって興奮が他のニューロンに伝わるには基礎網の中で樹状突起の分枝および神経突起(軸索突起)の分枝(図361)を介することもあり,あるいは神経終末足によって直接に細胞体およびその樹状突起の幹のところに伝わることもある(図358, 359, 362).これらのシナプシスは変りやすい構造であって,進行性および退行性の変化を示すのである(W. Kirsche, Psychiatr., Neurol., med. Psycholog., 6. Jahrg. ).
Bethe, A., Allgemeine Anatomie und Physiologie des Nervensystems. Leipzig, G. Thieme 1903.--Held, H., Monatsschr. Psych. NeuroL, 65. Bd.,1927,およびFottschr. naturw. Forsch., Heft 8,1929.--Boeke, I., Kon. Akad. Wetenschappen,1929. Bauer, Kar1, Z. Zellforsch., 30. Bd.,1940およびArch. Psychiatrie,114. Bd.,1941.
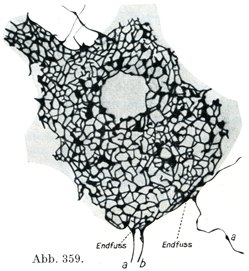
[図359]細胞周囲の神経終末網 イヌの脊髄前柱の細胞(Heldによる,1929).
4.一次,二次,さらに高次のニューロンNeuronen I., II. usw. Ordnung
各ニューロンは興奮を1つまたはそれ以上の場所から,1つまたはそれ以上の別の場所に伝えるのである.インパルス(刺激)Impulsはそこで直接に(神経原線維が1つのニューロンから他のニューロンに,中絶せずに移行しているとの仮定のもとに),あるいは間接に(興奮がたんに接触しているニューロンの間で伝わるという仮定のもとに)第2のニューロンに伝達してゆく.同じようにして第2のニューロンからインパルスが第3のニューロン,第4のニューロン等々に伝達される.いま問題としているこれらのニューロンは一次ニューロン,二次ニューロン,三次ニューロン等と名づけられている(図360~363).
ここで特に次のことを強調しておく,すなわち1個の低次のニューロンから興奮が多数のより高次のニューロンに伝わるが,また多数の相異なる場所からの興奮が単一のニューロンに伝達される.そのさい同一の興奮が中枢器官のいろいろ違った場所に達した後に,これらの場所からふたたび1個の神経細胞に達することがある.
Elze(Z. Anat. Entw.,94. Bd.,1931)は伝導弓Leitungsbögenと綜合場所Integrationsorteとを区別している.伝導路Leitungszvegeはみな末梢に始まり,中枢神経系を通ってふたたび末梢に帰る.伝導弓の頂点は中枢神経系の内部にあって,その求心性の脚と遠心性の脚とはその小部分だけが中枢神経系のなかにある,なぜならばそれより大きな部分は末梢神経系に属しているからである.直接反射弓direkter Reflexbogenのなかにあるような最も簡単な伝導弓はただ2つの脚,すなわち求心性のニューロンと遠心性のニューロンとだけからできている.第3のニューロン(脊髄の索細胞Strangzellen,脳の網様核Nucleus reticularis)をあいだに介して興奮が同側および反対側のかなり多数のニューロンに伝達される.これらの2節および3節からなる伝導弓を全部合せたものが脊髄および脳幹の固有装置Eigenapparatをなしている.
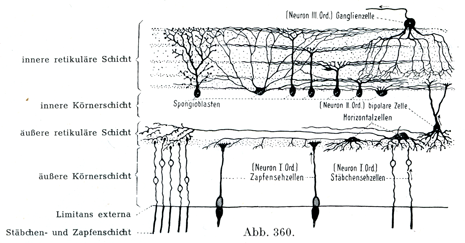
[図360]興奮が樹状突起および神経突起を介してニューロンのあいだを伝わることの例.クローム銀染色法による哺乳類の網膜(Cajalによる).矢印は神経伝導の方向を示す.
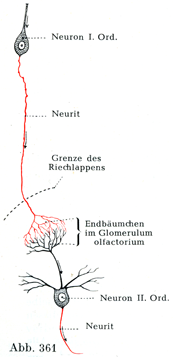
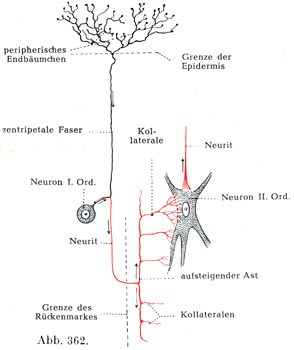
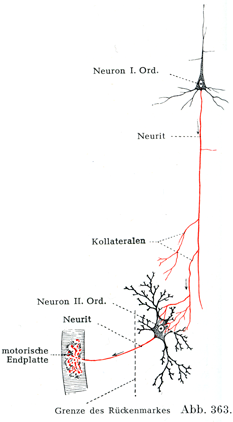
[図361~363]刺激が一次ニューロンから二次ニューロンに伝わる具合を示す模型図 矢印は興奮の伝ってゆく路を示す,神経突起は赤色.図361.一次ニューロンの終末分枝によって二次ニューロンの樹状突起の枝への刺激伝達が仲介されている.図362, 363.一次ニューロンの側枝の終末分枝によって二次ニューロンの細胞体への刺激伝達が仲介されている.
このような伝達弓は脳のあらゆる部分に存在する.かくして脳内の一定の場所(小脳皮質,中脳の視蓋,大脳皮質)は次のような特徴をもっている.すなわちこれらの場所は末梢の全体から求心性の脚が集まってくるが,しかしここからは(求心性の脚よりも)ずっと少い数のごくわずかな遠心性の伝導路だけが末梢にもどってゆくのである.それゆえこれらの場所では伝導路の集中が起るので,Elzeは綜合場所Integrationsorteと名づけている.この3つの綜合場所にわたっている伝導弓の全体が綜合装置Integrationsapparatを成すのである.
5.核と中枢Kerne und Zentren,伝導路と神経束Bahnen und Bündel,神経根と根線維Wurzelln und Wurzelfasern
同じ作用をいとなむニューロンの細胞体Zellkörperは概していっしょに相ならんで集まって群をなしている.その群は周囲に対してある程度境のついた区域をなしている.このような細胞体の群Zellkörper-Grappeはそれらの細胞の樹状突起および他のニューロンの終末分枝Telodendrenの混和した部分といっしょになっていて,中枢Zentrenあるいは核Nuclei, Kerneと呼ばれるのである.(脳神経のみに関しては)その細胞群が神経突起を末梢に送っているときは起始核Nuclei originisといい,求心性の伝導路がその細胞群に終わっているときは終止核Nuclei terminalesとよんで,両者を区別している.
それゆえ舌下神経核とは,その神経突起が集まって舌下神経を作るところのニューロンの細胞体群の全部をさし,言語中枢とはそこの細胞が言語を発することに特にはたらいている大脳皮質の一部をいうのである.
同じ機能をもつニューロンの細胞体と樹状突起だけがたがいに近くにあるのでなく,そういうニューロンの神経突起(軸索突起)Neuritenもやはり束をなして集り長短いろいろの差があるがたがいに並んで走っている.
神経突起のあつまつた比較的太い束は路Tracths, Bahnenあるいは束Fasciculi, Bündelと名づけられ,その細いものは線維束Faserzügeあるいは線維Fibrae, Fasern,糸Filaと呼ばれる.それぞれの伝導路を特徴づけるものは形態学的および機能的な性状である.そのはたらきによって知覚性sensibel(感覚性sensorisch)および運動性motorischの伝導路,ならびに連合路Assoziationsbahnen(Vetbindungsbahnen)を区別する.
中枢神経系に出入する神経突起束あるいは神経線維束は根Radices, Wurzelnと呼ばれる.また個々の神経突起を根線維Filum radiculare, Warxelfaserという.
6.ニューロンの分類Einteilung der Neuronen
神経突起が刺激を伝える方向により,またニューロンの位置によって,3つの大きな群が区別される,すなわち求心性,遠心性,および介在性のニューロンである.
1. 求心性ニューロンzentripetale Neuronenすなわち受容ニューロンrezeptorische Neuronenは末梢で受けとった刺激を中枢に伝え,ここでその刺激をさらに他のニューロンにあたえる.これらのニューロンは感覚器の神経(感覚神経sensorielle Nerven)であって,末梢では神経上皮細胞(嗅細胞,視細胞)の突起として始まるか,あるいは(単に知覚性のものとして)末梢の神経網あるいは特別な終末器(Gefühlsnervenのばあい)(Guttmann:Medizinische TerminologieによればGefühlsnervenはsensible Nerven(知覚神経)と同意語である.(小川鼎三))のなかで始まり,そのニューロンの細胞体は他の場所(脊髄神経節のなか)にある.
2. 遠心性ニューロンzentrifugale Neuronenすなわち奏効ニューロンeffektorische Neuronenはその神経突起によってインプルスが中枢から末梢の器官へと伝わる.これらのニューロンは横紋筋および平滑筋に達する,すなわち運動神経motorische Nervenである.平滑筋の神経に属してその重要な一部をなすものが脈管の筋肉に分布する神経,すなわち脈管神経Gefäßnervenであって,そのなかで脈管収縮神経線維vasokonstriktorische Fasern(血管運動神経vasomotorische Nerven)はその興奮が血管を収縮させるものであって,もう1つの脈管拡張神経線維vasoditatierende Fasernはその刺激によって脈管の能動的な拡張がひき起されるものである.刺激によって収縮が起きないで,運動を緩慢にさせたり(Verlangsamung)また停止(Stillstand)させるような運動神経は抑制神経Hemmungsnerwenとよばれる.生理学ではなおその他のはたらきをもつ脈管神経のあることが知られている.腺Drüsenに達して,そのはたらきに影響をあたえる遠心性神経は分泌神経sekretorische Nervenという.さらに特別な一種類の神経としてしばしば問題にされているのが栄養神経trophische Nervenと呼ばれるもので,これは組織の栄養Ernährungということに一定の影響をおよばすものとされているが,しかしその実在については疑問があり,おそらくこれは血管運動神経あるいは分泌神経に他ならぬものであろう.
3. 介在性ニューロンinterzentrale Neuronenは中枢器官のうちの異なる部分の間を結合するニューロンである.これらは決して一次ニューロンではなくて,いつもそれより高次のニューロンである.その神経突起の長さはすこぶるいろいろである.なぜならばこの神経突起は中枢器官のなかで隣り合せている部分を結びつけたり,あるいはたがいに遠く離れている部分を結合したりするのである.つまりこのニューロンのはたらきは連合ということにあるので,連合ニューロンAssoziations-Neuronenと呼ぼれる.
また中枢神経系以外でも末梢神経節のなかや末梢神経のなかには介在性ニューロンが存在する.
7.白質weiße Substanz,灰白質graue Substanz,膠様質gelatinöse Substanz
ニューロンの細胞体の部分が群をなして配列することおよび神経突起が集まって東をなす,その配列が核および伝導路の色合いの大きな差をおこすのである.有髄性の神経突起は白くて繻子のように輝いて見えるが,これに反して細胞体の集まっているところ,すなわち核は灰白色に見えて光沢がない.この差異は灰白質が豊富に血管をうけることによっていっそう著しくなるのである.これに反して伝導路は比較的に血管の乏しいことが有髄神経線維の白い輝きをいっそうはっきりさせるのである.
核集団Kernmasseと伝導路との間の,目立った色の違いはその場所がそれぞれきまっているので白質Substantia alba, weiße Substanxおよび灰白質Substantia grisea, graue Substanzと名づけるもとになった.グリヤが比較的多く集まっていると,その場所はいくぶん透明になっているので,そこは膠様質Substantia gelatinosaと呼ばれている.
神経細胞体に色素が含まれていることによって2, 3の場所では黒い色調,青い色調および黄色をおびた色調を生じ,黒核Nucleus niger(Nucleus nigerはNomina Anatomica JAponicaでは「黒核」と直訳されているが実用上は「黒質」と呼ぶことが多い.(小川鼎三)),青斑Locus caeruleus,灰白層Stratum cinereumという.
8.器官としての末梢神経の構造
末梢神経は神経線維,すなわち運動性ニューロンの神経突起(軸索突起)および知覚性ニューロンの樹状突起からなっている.神経線維は一部は有髄,一部は無髄である.脳脊髄神繹では有髄線維の方が多くて,交感神経では無髄線維が優勢である.それゆえ前者は白く繻子のように輝き,後者は灰白色である.個々の神経線維の太さはすこぶるいろいろであり,太い神経線維と細いものとが相並んでいる(図364).
個々の神経線維は結合組織によって分けられ,同時にまとめられている.結合組織は個々の神経線維の周りに薄い鞘,すなわち神経内膜鞘Endoneuralscheidenをなしている.この鞘は細い膠原線維と内皮細胞とからなるのである.多数の神経線維およびその神経内膜鞘がかなり幅のひろい結合組織束によってまとめられ細い1本の線維束となり,それをまとめる結合組織のなかには膠原線維のほかに弾性線維やかなり太い血管が含まれている.個々の神経線維のあいだにある組織の全体が神経内膜Endoneuriumとよばれるのである(図364).このような細い線維束のいくつかがいっそうかたい結合組織の数層をなしてたがいに同心性に配列しているもの(層板Lamellen)に取り巻かれており,この層板をなす結合組織の個々の束は主として神経の長軸の方向にのびている.これらの層板を神経周膜層板Perineurallamellenといい,その全体を指して神経周膜Perineuriumといい(図368).個々の神経周膜層板のあいだの隙間にはリンパ間隙Lymphspaltenがあり,内皮細胞が被っている.これらの隙間は中枢神経系を取り囲むところのリンパ腔Lymphräumeと続いている.
さらにいくつかの束が,血管および脂肪組織をもつ結合組織によって1つにまとめられている.この結合組織が神経上膜Epineuriumである.かなり太い神経では神経上膜の結合組織束は固く且つ丈夫であり,脂肪組織が豊富にあり,多数の血管がふくまれる.
神経の個々の束はその走行の途中で分岐する.その分れた枝が他の神経束に接して,しかもそれと合同している.こうして神経叢Nervenplexusが生ずる(それについては第10項, 295頁を参照せよ).
神経線維束と神経上膜の膠原線維とはNauck(Verh. anat. Ges.,1931)によれば波形を呈している.その波の形が延びてなくなることによって神経は伸長できるのである.
9.器官としての末梢神経節の構造
末梢の神経節Peripherische Ganglienは神経細胞が局所的に集まったものであって,多数の(有髄および無髄の)神経線維によって貫かれている.ごく少数しか細胞体を含まないで顕微鏡でやっと見えるような小さな神経節もある.
脊椎間の神経節および交感神経節の細胞の形Zellformenはいろいろである.脊椎間神経節の細胞はその大部分が単極であり(図366),交感神経節の細胞は多極である.
脊椎間神経節,すなわち脊髄神経節の細胞は学者によって,多少の差はあるが若干の数の型に分けられている.これを区別する目じるしとしては,細胞の大きさ,そのなかにあって染色のできる物質の量とその外観, 細胞の突起の数とその様子である.--また成人においてもなお成長しつつある細胞と考えられる胎生期の性質を有ったいくつかの細胞が,他の細胞とともに存在することも重要視されねばならない.
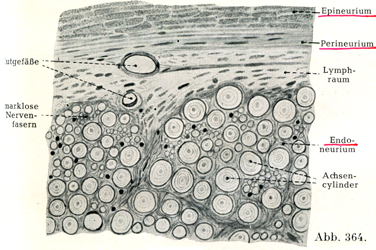
[図364]神経内膜,神経周膜,神経上膜;太い神経線維と細い神経線維.ヒトの坐骨神経の横断面の一部.
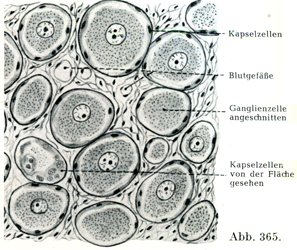
[図365]脊髄神経節の細胞とその結合組織性の被膜および(グリヤ性の)外套細胞 ヒトの脊髄神経節(C. VI)より.
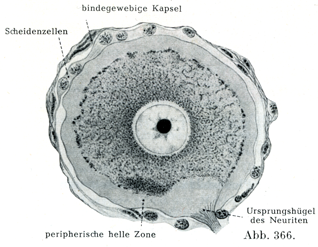
[図366]ヒトの脊髄神経節の大きな細胞(100µ)とその結合組織性被膜(Lenhossék)
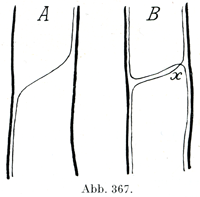
[図367]A. 単純結合Conjugatio simplex, B. 相互結合Conjugatio mutua,×印のところで交叉している.
神経節は全体として,厚薄いろいろな厚さの結合組織性の被膜に包まれている.これはそこにやってくる神経の神経周膜のつづきである.神経節の内容は神経線維および神経細胞である(図369).神経線維は神経内膜鞘に包まれており,神経細胞は殻の中にあるクルミのように特別な被膜の中にある.これらの被膜は神経線維の神経内膜鞘のつづきをなす結合組織線維と,なおまた被膜細胞Kapselzellen,外套細胞Mantelzellen,辺縁細胞Randxeltenあるいは鞘細胞Scheidenzellenと呼ばれるかなり多数の細胞とからできていて,この細胞は神経細胞の表面と線維性被膜の内壁との間にあり,Heldによれば末梢性のグリア細胞とみなすべきものである(図365, 366).
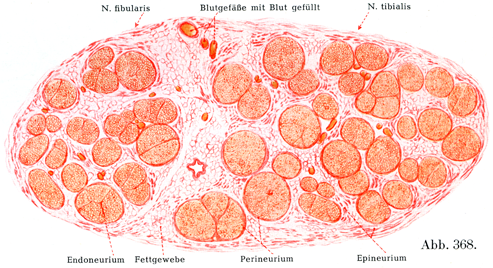
[図368]ヒトの坐骨神経 横断
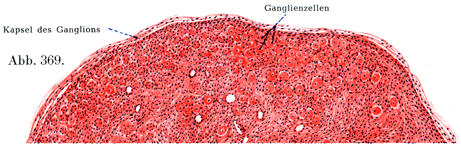
[図369]ヒトの下頚神経節 その一部を弱拡大で示す.
10.神経線維および神経の結合:神経叢形成Plexusbildung,神経の吻合Anastomosen
ニューロンの細胞体より発する神経突起は集合して束をなして,中枢器官の内部でも,また末梢神経系の中でもさらに遠くまでいっしょに走っている.そのさい個々の束が分れて,また他の束と合し,その全部あるいは一部がふたたびこの束から別れて,もう1つ別の束に加わったりする.この束の分れ且つ合する形式を神経叢形成Geflechtbildungといい,その結果できるものが神経叢Plexus, Geflechtである.
つまり神経叢というのは神経のあいだの複雑な形の結合である.これに対して単純な末梢牲の結合である吻合Anastomose,または結合Konjugationは太さのいろいろな末梢神経が別のそれと合することである.そのときにいくつかの特性が生じうる.すなわち--a)結合枝が1本の神経に属する線維を他の神経の路に送っている(図367, A).その線維が後者の中で中枢の方あるいは末梢の方に走っていることがあり,また両方向に走ることもある.この形を単純結合Conjugatio simplexと名づける.--b)その結合枝が両方の神経の線維を導いている.こうして相互結合Conjugatio mutuaが生ずる.その線維のすすむ方向がやはりあるいは中枢のがわに,あるいは末梢のがわに,あるいは両方向に進むものがあるわけで,その線維の列び方がいろいろ違ったものでありうる.その場合にはどこかある場所で交叉Decussationes, Kreuzungenが当然おこるはずである(図367, Bのxに示してある).そのような交叉は末梢神経系の中では幾度もみられるばかりでなく,中枢器官の中でもはなはだひろく存在する.これは特に中枢器官で体の両半に属する線維のあいだに正中面において起る重要な交叉についても同じことである.
これらの大きな中枢性の交叉の意義は,それが運動性のものでも,知覚性のものでも,すこぶる重大なものにちがいない.これらの交叉は,中枢性の交連Kommissuren と同じく体の両半が密接に一体をなすためのものであることを意味し,体の両半の中枢と末梢とはこれらの交叉によってたがいに深い交わりをするのである.
2-30

最終更新日 13/02/03