Rauber-Kopsch解剖学
歴史的な偉大な解剖学書
Rauber Kopsch Band2. 67 


7.尾骨神経叢Plexus coccygicus, Steißbeingeflecht(図565)
これは小さい神経叢であって,SVとCoIとの一部よりなる.上方では(仙尾骨神経係蹄Ansa sacrococcygicaにより)陰部神経叢とつながる.いく本かの短い小枝が尾骨神経叢と交感神経幹の下端部,すなわち第4あるいは第5仙骨神経節および尾骨神経節とのあいだを結びつけている.
仙尾骨神経係蹄から,あるいはSVそのものから肛門尾骨神経N. anococcygicusがでている.これは尾骨筋の前面の上を走り,次いでこの筋と肛門挙筋とのあいだをへて背側面に入り,尾骨角の外側で皮膚の下に達する.ここで尾骨神経の後枝Ramus dorsalisからの1小枝と結合して,若干の小枝(すなわち肛門尾骨神経Nn. anococcygici)を肛門と尾骨のあいだの皮膚にだし,また尾骨筋神経N. coccygicusに属する諸枝が後方に走り,尾骨じしんの後面を被う皮膚に分布している.
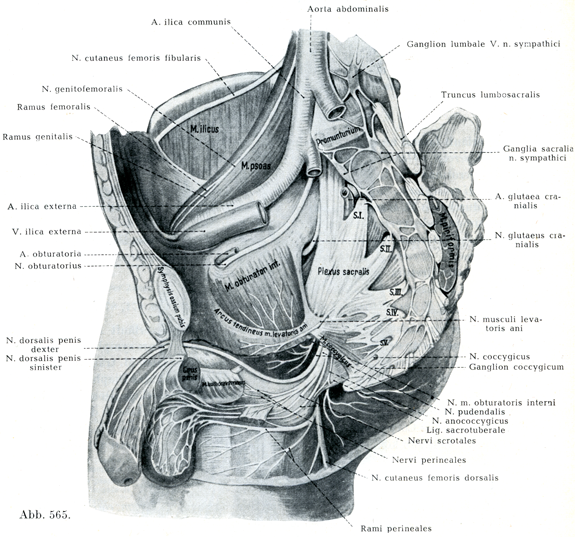
[図565]男の骨盤の右半の内面および諸神経の分枝(3/10) (HirschfeldおよびLeveilléによる)
D. 交通枝 Rami communicantes(図371, 530, 570)
胸神経の領域にある交通枝が最もかんたんな様子を示している.そこでは交通枝が後枝のおこるところに向き合った個所,あるいはそのそばですぐ前方のところで胸神経からでて,ついで鋭角をなして内側,下方,かつ前方にすすみ,近くにある[交感]幹神経節の外側縁に入る.神経節の上枝Ramus cranialisあるいは下枝Ramus caudalisに入ることは極めてまれである.
交通枝は1本であるか,あるいは2本の小枝に分れており,ときには3本に分れ,これらの小枝はたがいに平行に列んでいるか,あるいはかなりの間隔をおいて脊髄神経の幹から発して1つの[交感]幹神経節に向かって集中している.これに反して1つの交通枝に属する小枝が2つの異なる[交感]幹神経節に入るばあいはたがいに遠ざかつている.
頚神経の交通枝はその数においても,結合のしかたにおいても胸神経のばあいよりもいっそう大きい変化を示すのである.この変化は本質的には頚部の交感神経節の長さがいろいろと異なることにより,また中頚神経節が欠けることによってひきおこされる.これは第1と第2頚神経の交通枝,第7と第8頚神経の交通枝が頚部では最も規則正しい配列を示すことと連関がある.
第1頚神経の交通枝は直接にその前枝から,あるいは第1頚神経係蹄Ansa cervicalis primaから,あるいはその前枝と舌下神経とが作る係蹄からおこる.第2頚神経の交通枝はその前枝から出るのがふつうである.第1と第2頚神経の交通枝は上頚神経節に達している.
第3と第4頚神経の交通枝は直接にその前枝から発することもあり,あるいはそれらの頚神経がつくる係蹄から出ることもある.これらの交通枝は深頚筋群の上(前方)あるいは下(後方)を通って交感神経幹に達する.第3頚神経の交通枝もやはり上頚神経節に加わる.この神経節は,いくつかの神経節,それは少くとも4個の神経節が融合して1つになったものである.第4頚神経の交通枝も第3のものと同じぐあいになっているか,あるいは第5と第6頚神経の交通枝と同じく中頚神経節が存在するときにはこれに入っている.あるいはまた第4頚神経の交通枝は第5と第6頚神経の交通枝がやはりそうであるように,中頚神経節が欠けているときには上頚神経節と下頚神経節とのあいだにあるこれら両神経節の結合索Verbindungsstrangに接着するのである.
第7と第8頚神経の交通枝は交感神経の下頚神経節に達する.第6交通枝が同じ関係を示すばあいもすでに知られている.
腰神経の交通枝は長くて,これらは椎間孔から腰椎の前面までの長い道のりをすすむのである.これらの交通枝はこの長い道を横あるいは軽く斜めの方向にすすみ,そのさい大腰筋の筋束のあいだを走り,ついで椎体の前面と大腰筋の内側の腱弓とのあいだで上下方向にのびている裂け目のなかに現われる.これらの交通枝はふつう重複しており,しばしば次のように配列している.それは同一の神経節が2つの異なる腰神経と結合する,しかしまた別のばあいは1つの交通枝が小枝に分れて,2つの隣接する神経節と合している.
最後に述べたことはまた仙骨神経の交通枝についてもあてはまる.仙骨神経の交通枝はしばしば重複しているが常に短くて,前枝が前仙骨孔から出るところですぐにこれから分れて,内側に向い,近くの[交感]幹神経節に達し,そのさいこれらの交通枝は外側仙骨動脈の上を越えるのである.
上に述べた数多くの交通枝が脊髄神経系あるいは脳脊髄神経系と交感神経系とのあいだを結びつける.脳神経にも交通枝は豊富に存在する.交通枝はふつうに交感神経性の根Wurzelnと名づけられている.根という名前は交感神経幹に脳脊髄神経系の線維を導くということを意味している.交感神経幹には交通枝によって効果性ならびに受容性の線維が導かれるのである.この両種の線維は脳脊髄神経の両根に由来している.しかしまた交通枝によって脳脊髄神経系に交感神経性の線維が導かれる.それゆえ交通枝は一部は脳脊髄神経性の枝,一部は交感神経の枝であり,後者の中心は交感性の神経節のなかにある.
交感神経の線維が交通枝によって脊髄神経の末梢部に導かれ,それも主としてその前枝Ramus ventralisの末梢部に導かれる.前枝が脊髄神経からでる4つの枝のうちで最も太い枝でありその分布領域も最も広い.しかしまた後枝Ramus dorsalisおよび硬膜枝Ramus meningicusのなかにも交感性の線維が達してい為. 最後にとりわけ多量の交感神経の線維が交通枝Ramus communicansのあらゆる末梢分布にしたがって内臓と血管に達するのである.
身体の多くの場所では上述のように各脊髄神経への交通枝が2本あるいはまた3本も存在する.これらのうちで1本嫡主として髄鞘をもつ脊髄性の神経線維より成り,他の1本は特に交感神経からの灰白線維(無髄)を有っている.そのばあい前者は白く見え,後者は灰白色に見える.しかしたいていは両交通枝のそれぞれに両種の線維が混在している.
脳脊髄神経とその神経節の構造
a)神経幹と神経根Nervenstämme und Nervenwurzeln
脳脊髄神経は主としてシュワン鞘 Schwannsche Scheideをもった有髄性の線維よりなり,反射光のなかで白く見える.この有髄神経線維には一部は散在した状態で,また一部は小さい束にまとめられて無髄神経線維がまじっている.脳脊髄神経の束は豊富な結合組織によって包まれ,かつこれによって貫ぬかれ,この結合組織は特別なぐあいに配列している.神経根では結合組織が初めは柔膜の続きどしてわずかに存在するのみであるが,神経根が外に出てゆくにつれそれがだんだんと豊富になり,そのさいクモ膜鞘と硬膜鞘とがそれぞれの髄膜の続きとしてこれらの神経根を包み接着している.
かくしてこれらの神経ははじめは脊髄および脳と同じ被膜をもっているが,脊髄神経節の前でこれら3つの鞘がたがいに豊富なつながりをなして合するのである.硬膜はますます疎になり,脂肪組織をふくんで,外方へも内方へも境がはっきりしなくなる.
末梢神経のなかでの神経線維と結合組織との配列については294頁を参照されたい.ここでは次の点だけを付け加えておく:
神経上膜と神経周膜鞘の厚さは末梢にゆくほど次第に薄くなり,そのうちに個々の線維束は早かれ遅かれいずれは結合状態から離れてゆく.そうなると個々の線維束は多くのばあいただ1つの神経周膜鞘をもつのである.最後に小さい神経幹から別々に枝分れしてでる神経線維Nervenfasernはなお神経周膜板の薄い内皮性の続きによって包まれている.この被膜をRanvierはヘンレ鞘Henlesche Seheideと名づけた.このヘンレ鞘とシュワン鞘とのあいだにある狭い腔所は液体を注入することのできるリンパ腔Lymphraumであって,神経のその他のリンパ系と直接につながっている.
末梢神経の血管Blutgefäßeである小動脈と小静脈とは神経の走る方向に沿って,まず神経上膜のなかを進んでいる.さらに数多くの細い血管が神経周膜鞘を貫いて2次線維束の内部に入る.そこではこれらの細い血管が長く延びた網目構造のきれいな毛細管網に移行する.これらの小動脈は細い血管神経Gefäßnerven(Nervi nervorum「神経のもつ神経」の意)を伴っている.
神経根Nervenwurzelnは中枢に向かっては脊髄に入るところまでシュワン鞘をもっている.脊髄ではシュワン鞘の代りにグリアNeurogliaがある.
b)脊髄神経節(図399, 566)
脊髄神経節はその主要な構成要素として神経細胞と神経線維とをもっている.なおこれに加えて結合組織,血管およびリンパ管がある.その神経細胞は大部分が偽単極性のものである.すなわちただ1本だけ存在するその突起がなお神経節の内部にあるあいだたT字形,あるいはY字形に中枢がわと末梢がわとの2本の枝に分れる.これらの神経細胞のほかには もっともその数はずっと少ないのであるが一なお別の3種類の細胞がある:その1つは小さい細胞でただ1本の突起をだす.この突起は脊髄神経に達するが,それからおこる側枝は神経節のなかにとどまるのである.他のものは小および中等大のいわゆる介在細胞“Schaltzellen”であって,それらがもついくつかの突起はこの神経節の範囲を越えて外にはでないのである.また最後のものはすでに上に(295頁)述べた胎生期の性質を示す細胞ならびにそれから発達しつつある細胞である.
脊髄神経節の細胞の微細構造については294~296頁,図365, 366ならびに第I巻では図120,124を参照せよ.
脊髄神経節における線維の走り方および脊髄と交感神経幹との結合はHirtによれば次の如くである(図399):
相異なる6つの型を脊髄神経節のなかのニューロンに区別することができる.第1~第4のものは偽単極性の細胞,第5のものは単極性の細胞,第6のものは多極性の細胞である.
1. 定型的な体知覚性の細胞“somatosensible Zellen”,その末梢がわの突起は末梢のどこかから発し,その中心がわの突起は後根線維として脊髄に達して,ここで以前(313, 316, 320, 322頁)述べたぐあいに終わっている.
2. 内臓知覚性の細胞“viscerosensible Zellen”,その末梢がわの突起は交感神経幹を介して植物性神経の支配をうける何れかの器官からやって来る.その中枢がわの突起は後根線維として脊髄に達し,そこで体知覚性の細胞と全く同じ結合をなし得るのである.
3.2本の突起とも求細胞性cellulipetalに導く細胞:その末梢がわの突起は前者と同じく末梢のどこかからはじまるのであるが,中枢がわの突起は後根の内部で終わっている.
このニューロンは知覚性の興奮を末梢および後根から脊髄神経節に導くのであって,ここでは介在細胞によって別の細胞に伝達するようになっている.
4.2本の突起とも遠細胞性celtulifugatに伝える細胞:その中枢がわの突起は後根線維として脊髄に入り,側柱の細胞群に達する,それゆえ求心性zentripetalであるが,末梢がわの突起は遠心性に伝える]のである.
これらの細胞は体知覚性の細胞および内臓知覚性の細胞から介在細胞を介して,あるいは交感神経からの線維によって興奮を受けとる.
5. 単極性の細胞ecnipolare Zellen,その突起は脊髄神経の内部に,あるいは交通枝を通って交感神経に達し,両方のばあいとも血管に終わっている.
これらは脊髄神経節のもつ血管運動性の細胞である.別の説によれば血管収縮性の(交感神経性の)線維Vasoconstrictorenは前根を通り,血管拡張性の(副交感神経性の)線維Vasodilatatorenは後根をとおるという.
6. 介在細胞“Schaltzellen”,これは多極性の細胞であって,そのいく本かの突起がこの神経節の範囲にとどまり,それを越えて外にでることはない.それゆえただ神経節内の結合のはたらきをしているに過ぎない.
そのほかにいわゆる通過線維“durchtretende Fasern”が神経節を貫いて通りぬけている.
脊髄神経節の結合線維は神経根のそれと同じく髄膜の続きをなすものと見なされる.個々の細胞とそれからでる線維はそれぞれ結合組織性の鞘の続きによって包まれている(図365, 366参照).
神経の血管Blutgefäße der Nerven
Tonkoffによれば神経栄養動脈Arteria nutricia nervorumと神経伴行動脈Arteria comes nervorumとが区別されなければならない.後者は皮膚の動脈と筋肉の動脈とのあいだを吻合させるものであり,側副路の形成にさいして重要なものとなり得る.
第1頚神経節はその栄養枝Ramus nutriensを直接に椎骨動脈から受ける.すべての脊髄神経節はその動脈をそれぞれ2つまたはそれ以上の源から受けとるのである.最も重要な源をなすのは体分節性の動脈の脊髄枝Ramus spinalisから出るRamulus medius(中央の小枝)であって,これはA. radicalis medullae spinalis(脊髄根動脈)といおれるものに続いている.Tonkoffはすべての太い神経幹についても正確な報告をなしている(Internat. Monatsschrift f. Anat. und Phys.,15. Bd.,1898).
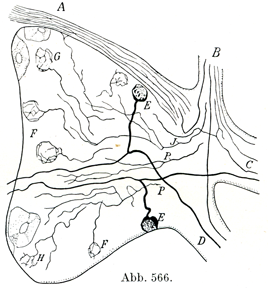
[図566]生後10日のラットの脊髄神経節1個の横断
クローム銀染色(Cajal) この図には細胞周囲の定型的な分枝をいろいろな標本からまとめ合せて画いてある.
F, G, H細胞周囲の神経線維籠Nervenfaserkörbe;P, J 神経節のなかで枝分 れする線維;A前根;B交感神経根(交通枝);C 脊髄神経の前枝;D 脊髄神経の後枝;E 脊髄神経節の細胞.
神経分節・筋分節および皮膚分節の互いのあいだの関係
節をなしてつぎつぎに並ぶということは周知のごとく神経系Nervensystemと血管のみに見られるのではなく,さらにまた,それもいっそうはっきりした程度で筋肉Muskulaturと骨Knochenがそうである.なおまた,全身の皮膚Integumentすなわち外皮が神経と血管の支配によって分節に関与している,つまり皮膚分節Dermatomeren, dermale Segmenteが分けられるのである.また腸もそれ以外の体に共通する配列を示さないとはいえない.もっとも腸では,特にその腹部にある部分ではすでに早期に小腸と大腸の長さが著しく増すことによって,分節の境が完全になくなってしまうのである.しかし腸およびその他の内臓のすべてをみると,その神経および」血管の分布する関係はその発生史と相まって腸分節Enteromerenの厳存することを示すのである.
これらすべての分節構造を生ずることについての最も直接的な誘発要素はおそらくは筋肉Muskeclaturが かなり早期に分節的な配列を示すことに求めるべきであろう.
各神経分節にはそれぞれ相応した筋分節,皮膚分節,腸分節が属している.その発達のあいだに同一の体分節に属する神経,筋などの分節は高度にその位置が移りかわるので,1つの分節に属する諸部分を知ることは骨の折れる研究を必要とする.とくに皮膚分節および筋分節とそれに相当した神経分節との関係を知ることは実地医学において大きな意義がある.
それゆえ次にこの関係を1つの表にまとめてみよう.この表では神経分節とそれに属する筋領域と皮膚領域との関係が示され,また反射に関係する諸分節も示してある.
脊髄のいろいろな分節における機能の局在
StarrとEdingerが動物実験と病理学的な観察とに基づいてまとめたものによる.個々の筋について少しばかり異なる報告がされているが,それをこの表と比較されたい(第1巻).
|
分節 |
筋 |
反射 |
皮膚の感覚支配 |
|
|
第2~第3頚分節 |
胸鎖乳突筋の胸骨乳突部 |
肋骨弓に急激に圧を加えたときの吸気 |
項部および後頭部 |
|
|
僧帽筋 |
||||
|
斜角筋群および項部の諸筋 |
||||
|
横隔膜 |
||||
|
第4頚分節 |
横隔膜 |
項部の刺激による瞳孔散大,第4から第7までの頚分節 |
項部 |
|
|
棘上筋と棘下筋 |
肩の上部 |
|||
|
三角筋 |
上肢の外方面 |
|||
|
上腕二頭筋と烏口腕筋 |
||||
|
腕橈骨筋 |
||||
|
菱形筋群 |
||||
|
第5頚分節 |
三角筋 |
肩甲反射,第5頚分節から第1胸分節まで |
肩および上肢の後面 |
|
|
上腕二頭筋と烏口腕筋 |
相当する筋の腱反射 |
|||
|
腕橈骨筋と回外筋 |
上腕および前腕の外方面 |
|||
|
大胸筋の鎖骨部 |
||||
|
外側鋸筋 |
||||
|
菱形筋 |
||||
|
上腕筋 |
||||
|
小円筋 |
||||
|
第6頚分節 |
上腕二頭筋 |
上腕および前腕の伸筋群の腱反射 |
前腕の外方面 |
|
|
上腕筋 |
||||
|
大胸筋の鎖骨部 |
||||
|
外側鋸筋 |
||||
|
上腕三頭筋 |
||||
|
手および指の伸筋群 |
手関節の腱第6から第8までの頚分節 |
手背面,橈骨神経の分布領域 |
||
|
回内筋群 |
||||
|
第7頚分節 |
上腕三頭筋の長頭 |
手掌を叩くと諸指を閉じること |
橈骨神経の分布する手の領域分節 |
|
|
手と指の伸筋群 |
||||
|
手の屈筋群 |
||||
|
手の回内筋群 |
||||
|
大胸筋の胸肋部 |
|
分節 |
筋 |
反射 |
皮膚の感覚支配 |
|
|
肩甲下筋 |
手掌反射,第7頚分節から第1胸分節まで |
|||
|
広背筋 |
正中神経の分布領域 |
|||
|
大円筋 |
||||
|
第8頚分節 |
手と指との屈筋群 |
瞳孔反射 |
尺骨神経の分布領域 |
|
|
手の小筋群 |
||||
|
第1胸分節 |
母指の諸伸筋 |
|||
|
手の小筋群 |
||||
|
母指球と小指球 |
||||
|
第2~第12胸分節 |
背部と腹部との諸筋 |
腹壁反射,上腹部・第4~第7胸分節 |
胸部・背部・腹部・上臀部の皮膚 |
|
|
背部の諸伸筋(固有背筋) |
腹壁反射,腹部,第7~第11胸分節 |
|||
|
第1腰分節 |
腸腰筋 |
挙睾筋反射第1~第3腰分節 |
恥部の皮膚 |
|
|
縫工筋 |
陰嚢の前面 |
|||
|
腹筋群 |
||||
|
第2腰分節 |
腸腰筋 |
膝蓋腱反射,第2~第4腰分節 |
寛骨部の外方面 |
|
|
縫工筋 |
||||
|
膝関節の諸屈 |
||||
|
筋(Remak?) |
||||
|
大腿四頭筋 |
||||
|
第3腰分節 |
大腿四頭筋 |
寛骨部の前面および内方面 |
||
|
大腿の内回旋筋群 |
||||
|
大腿の内転筋群 |
||||
|
第4腰分節 |
大腿の内転筋群 |
寛骨部と下肢の内方面で脛骨躁までと足の内方面 |
||
|
前脛骨筋 |
臀筋反射,第4~第5腰分節 |
|||
|
膝関節の諸屈 |
||||
|
筋(Ferrier?) |
||||
|
第5腰分節 |
股関節の外回旋筋群 |
|||
|
膝関節の諸屈筋(Ferrier?) |
||||
|
足の諸屈筋 |
寛骨部の後面,大腿の後面,足の外側部 |
|||
|
足の指の諸伸筋 |
||||
|
腓骨筋群 |
||||
|
第1と第2仙骨分節 |
足とその指との諸屈筋 |
足底反射 |
大腿の後面,下肢と足との外側面 |
|
|
腓骨筋群 |
||||
|
足の小筋群 |
||||
|
第3~第5仙骨分節 |
会陰の諸筋 |
アキレス腱反射 |
仙骨を被う皮膚,肛門,会陰,外陰部 |
|
|
膀胱および直腸反射の中枢 |
神経が筋に入るところとその中での分枝
von BardelebenおよびFrohseの研究をもとにして次のことを述べておこう:神経が入る場所とその神経の分枝とは筋の形に相応していることもあるが,いつもそうとは限らない.それぞれの神経は同じくらいの太さの2本の枝に分れるか,あるいはその幹が順順に枝を1本ずつ出している.また筋に分布する神経のどれもが血管神経を出している.神経が血管と伴って筋に入ることもあり,あるいは別々の所で入っている.そしてまた各神経が逆行する枝をそれぞれ1本出している.
その入る場所は筋の深い方の面,あるいは表ての方の面,そして近位部,あるいは筋の近位端から1/3のところ,または幾何学的な中点(これはまれである)にあって,決してはなはだしく遠位のところに入ることはない! 筋の中における神経の分枝のしかたにはすこぶるいろいろな形がある:大部分が下行する核であったり,長い下行枝と短い上行枝,同じ長さの上行枝と下行枝のことがあり,枝が扇形に放散していること,円錐形に放散していること,1つのがわあるいは2つのがわに向かって終末小枝の形で枝分れすることもある(第I巻,340頁をも参照せよ).
筋外と筋内の係蹄,また筋外と筋内の結合があり,筋内神経叢がある.筋内神経叢はEislerによれば各筋によって全く一定した1つめ型を示している.
Köllikerによれば筋線維の長さが神経の関係において主要な役目を演じている.1つの筋において筋線維が筋じしんと同じだけの長さであると,神経はその筋の中では1つの場所に限つて広がっている.
縫工筋の各筋線維はそれぞれ3~4個の運動終板を有っていう(Sandmann).このことから個々の筋線維がもつ運動終板の数も考慮に値いする.
筋の神経分布を理解するのに最後に重要なことは筋の発生のことであり,それも個体発生と系統発生の両方である.
嗅糸Fila olfactoriaは第1脳神経をなしており,その由来については脳神経と脊髄神経に属する知覚性の根の発生とある程度の関係を示すのである.
終神経N. terminalisは明かに嗅糸と近い関係にある.
視神経Fasciculus opticus, Pedunculus opticus v. Wijhe, Augenstielはその他の脳神経や脊髄神経と形態学的な関係をもつような末梢神経では決してなく,脳の異なる部分を結びつける中枢部間の1つの結合索である.それが結合している脳の部分というのは一方では網膜,他方では四丘体と間脳と終脳である.
第3と第4と第5の脳神経は動眼神経N. oculomotorius,滑車神経N. trochlearis,三叉神経N. trigeminusであり,これらに三叉神経群Trigminus Gruppeとして総括することができる.
この三叉神経群の中では三叉神経じしんが第1鰓弓erstei Kiemenbogen,すなわち顎骨弓Kieferbogenの神経である.それは鰓分節Brdhchiomerie,すなわち腸壁と体壁前方部の分節構造に判断の標準をおいて,また顎骨弓から出来てくる諸構造の神経支配を考慮すると,こうなるのである.
そうすると三叉神経の第1枝は鰓前節様構造Präbranchialer Metamerに属している.動眼神軽と滑車神経とは三叉神経の領域に属する.特にそれらの起始核を考えてみると,これは運動性起始核の内側部にあたっており,一方では三叉神経の運動核がその外側部をなしている.
内耳神経-顔面神経は第2鰓弓の神経であって,ここでもまた分類の基礎として鰓分節Branchiomerieが役だつのである.内耳神経はその形態学的な本質からは,聴覚および大きな平衡装置の任務をもつにいたった1つの知覚性の皮神経である.この群には運動性の内側の神経として,外転神経N. abducensが属している.この神経の起始核のある位置がそれに相当している.
第9と第10と第11脳神経,すなわち舌咽神経N. glossopharyngicus,迷走神経N. vagus,副神経N. accessoriusはいわゆう迷走神経群Vagusgrmppeをなしている.
舌咽神経Nglossopharyngicusは第3鰓弓の神経である.
しかし迷走神経N. vagusはそれより下位のいくつかの鯉弓の神経であり,これらの鯉弓に属する神経がまとまって1つの強大な総合神経をなしている.すなわち迷走神経と舌咽神経とは神経幹のなかにある神経節をそれぞれ2つもっていて,われわれが今日までに知りえた限りでは,これらの神経節はすべてたがいに一致した定まった構造を有つのである.
副神経N. accessoriusは脊椎動物のかなり高等なものではじめて独立レた構造物として現れるもので,これは迷走神経根の運動性の部分,すなわち疑核Nucleus ambiguusにはじまるものと同一の系列に属するのである.
第12脳神経,すなわち舌下神経N. hypoglossusは部分的に妹舌咽神経と迷走神経の分布範囲よりももっと前方の領域に分布するのであって,すでに古くJ. Müllerが想像したようにかなり多くの脊髄神経に相当し,これらには多くは(知覚性の)後根を欠いている.これらの脊髄神経がかたまってただ1つの舌下神経となり,それが2次的に頭の領域に入って,脳神経の1つとなったものである.
この考え方はFroriepによって確証された,彼は初めて(胎児において)舌下神経に属する知覚性の神経節を見いだしたのである.
2-67

最終更新日 13/02/03