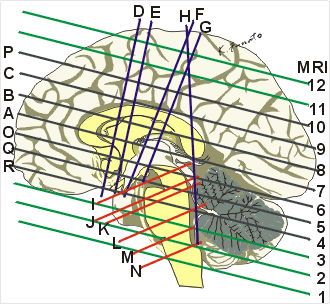
30歳♂のT1強調MRI画像
外眼角耳孔面に水平な面(島、線条体、内包と視床が見える) Aに対する面
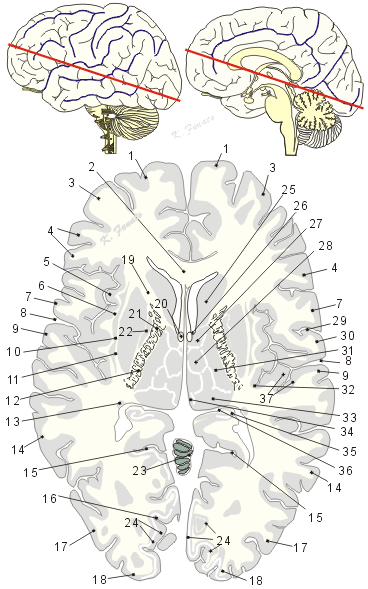
01:上前頭回、02:脳梁、03:中前頭回、04:下前頭回、05:島、島葉、06:最外包、07:中心前回、08:外側溝後枝、09:上側頭回、10:前障、11:外包、12:内包後脚、13:尾状核尾、14:中側頭回、15:頭頂後頭溝、16:鳥距溝、17:後頭葉の大脳回、18: 後頭極、19:内包前脚、20:脳弓、21:内包膝、22:淡蒼球、23: 小脳虫部、24:視覚皮質、有線野、25:尾状核頭、26: 室間孔、モンロー孔、27:視床前核、28:視床内側核、29:中心溝、30:中心後回、31:視床後外側核、32:島輪状溝、33:視床枕核、34:手網核、35:海馬、36:海馬采、37.横側頭回
01. Gyrus frontalis superior(上前頭回)Superior frontal gyrus
→上前頭回は上前頭溝の上にある。上前頭回および中前頭回の後部には運動性皮質中枢の続き(運動前野)があるが、これらの回の前部および下前頭回は連合中枢(前頭連合野)と考えられれる。
02. Corpus callosum(脳梁)Corpus callosum
→脳梁は左右の大脳皮質、ことに新皮質を結合する線維の集合したもので、系統発生的には最も新しく、ヒトでは非常に発育がよい。その前後経はほぼ7.7cmである。脳梁は正中断では全体としては釣針状で、4つの部分が区別される。後端部は膨大し、脳梁膨大といい、その前方に続いて水平に走る部分を脳梁幹とよぶ脳梁はその前端では強く屈曲し、脳梁膝をつくる。これはさらに後下方にくちばしのように尖って脳梁吻となり、しだいに薄くなって終板に続く。
03. Gyrus frontalis medius(中前頭回)Middle frontal gyrus
→中前頭回は上前頭溝と下前頭溝の間にあり、中前頭回および下前頭回は人脳では特に発育が良い。
04. Gyrus frontalis inferior(下前頭回)Inferior frontal gyrus
→下前頭回は外側溝の前枝と上行枝によって3つの部分に分けられる。すなわち、①眼窩部、②三角部、③弁蓋部である。優位大脳半球(通常は、右利きの人では左側にある)の三角部と弁蓋部は運動性言語中枢motor speech center(ブローカ中枢Broca's area)があり、言語活動に必要な微妙な運動支配を支配すると言われる。前頭葉下面は前頭骨眼窩部の上面にのっておりわずかに凹んでいる。ブローカ中枢は、右利きの人では左半球に、左利きの人では右半球にあるという説がある。
05. Insula; Lobus insularis(島、島葉)Insula Insula; Insular lobe
→ライルの島とも呼ばれる。外側溝の深部にある大脳皮質で、その表面は前頭葉、頭頂葉および側頭葉によりおおわれている。島をおおっているこれらの大脳葉の部分を前頭弁蓋、前頭頭頂弁蓋、および側頭弁蓋という。すなわち、弁蓋は島をおおう外套部である。島の周囲は輪状溝により囲まれ、これにより弁蓋と境される。この溝は島の下端部では欠き、この部分を島限とよぶ。ほぼ胎生17週からこの部はその周囲が厚くなるため、陥没してその輪郭が明瞭になり、第19週ごろから前頭葉、側頭葉および頭頂葉の発達につれてしだいにこれらによっておおわれ、生後は全くかくれてしまう。このように島の表面には前頭葉、頭頂葉、および側頭葉の部分が延びて来て、外側溝後枝の上下唇をなし、島を被っている。島は後上方から前下方に走る島中心溝によって後方の1~2個の長回と前方の4~5個の短回に分かたれる。Reil, Johann Christian (1759-1813)オランダ人解剖学者。精神病理学者。大脳のライル島を記述("Exercitationum anatomicarum fasciculus primus.etc", 1796)、生体の生理学機能の、化学的表現としての生命力を提唱("Von der Lebenskraft", Arch. Physiol, (Halle), 1796, 1,8-162)。最初の生理学雑誌"Arch. Physiol."と最初の精神病学雑誌"Magazin fur Nerven heilkundle"を刊行。
06. Capsula extrema(最外包)Extreme capsule
→前障の外側方には幅の狭い線維層があって、島の皮質と前障を隔てている。これらの線維の連絡関係は知られていない。
07. Gyrus precentralis(中心前回)Precentral gyrus
→大脳半球の外側面で、中心溝のすぐ前にある高まりが中心前回で、ここは随意運動に関係する運動領(運動野)motor areaである。
08. Rami posterior, sulci lateralis cerebri(外側溝後枝)Posterior branch of lateral sulcus
→外側溝の縁上回へ終わる長い枝。
09. Gyrus temporalis superior(上側頭回)Superior temporal gyrus
→上側頭回は外側溝と上側頭溝の間にある。上側頭回の後部付近には感覚性言語中枢sensory speech center(ウェルニッケ中枢Wernicke's area)があり、これは聞いた言葉を理解する中枢であるという。一次聴覚野は聴覚の中枢で、側頭葉の上側頭回の上面(Brodmannの41野)にある。二次聴覚野は一次聴覚野の周囲(42、22野)にある。一次聴覚野で聞く音の意味はこの皮質領域で理解される。ウェルニッケ中枢の障害時には、言語の理解ができない。ちょうどしらない外国語を聞くのに似ている。また、自己の発する言語音を聞きながら発声をすることができないので、理解できるような言語を発することもできなくなる。乳児のようにチンプンカンプンの発語となる(jargon aphasia)。このように感覚性言語野の障害で起こる失語を感覚性失語症sensory aphasiaまたは言語聾word deafnessという。感覚性言語中枢と聴覚領のすぐ下の上側側頭溝には、眼で見た物の動きを総合して認識する神経細胞が存在する。一側の一次聴覚野の障害では、反対側の聴力に障害が起こるが、聴力が完全に失われることはない。一側の聴覚器からの入力は両側の聴覚野に達するためである。二次聴覚野の障害が起こると、聞く音の意味を理解することができない。これを聴覚失認auditory agnosiaという。
10. Claustrum(前障)Claustrum
→前障はレンズ核と島との間にある、内側が凹面をなす板状の核で、腹側方に厚くなる。この核とレンズ核との間には外包があり、また島の皮質との間には最外包がある。これらは狭い白質で、大部分は連合線維から、一部は交連および投射線維からなる。前障は種々の視床核、扁桃体などから線維を受け、大脳皮質に広く投射する。前障は以前は線条体とともにいわゆる基底核に数えられたり、あるいは皮質層の付け足しとして島皮質に属するものとされた。しかしながら、発生学的ならびに比較解剖学的研究によって、前障は発生の途中で位置がずれた古皮質の細胞群であることが証明されている。前障はその広い底の所で古皮質の領域へ移行する(すなわち梨状前野や扁桃体の外側核へ)。頭頂葉、側頭葉および後頭葉の皮質からの、無髄線維が局在的配列をなして前障に終わると言われている。前障の機能についてはわかっていない。
11. Capsula externa(外包)External capsule
→前障と被殻の外表面とのあいだには幅の狭い白質の層があり、大脳皮質から被殻に達する神経線維はこの線維層を通る。
12. Crus posterius, capsulae internae(内包後脚)Posterior limb of internal capsule
→内包後脚の主部は大部分視床とレンズ核の間の空間を占め、ほぼ垂直の方向に走る線維群からなる。その前部に皮質脊髄線維(頭部を除く上半身に対する線維は前、下半身に対するものは後)があり、皮質脊髄線維の外側を皮質赤核路が走る。皮質脊髄線維の後方には上視床脚(視床後腹側核から大脳皮質の知覚野に至る線維および前外側腹側核から運動野と運動前野に行く線維を含む)がある。その後方に、レンズ核の後方をほぼ水平に後頭葉に向かう部分がある。これは後脚のレンズ後部で後視床脚、視放線、頭頂橋路、皮質被蓋路などがここを通る。後脚のレンズ下部はレンズ核の腹側を外側方に延びて側頭葉に向かう部分で、下視床脚、視放線および側頭橋路を含む。系統発生学的に古い投射路は古皮質および原始皮質から起こり、通常内包を通らず皮質かの脳部に至るもので、そのうち特異な所見を示すのは脳弓である。
13. Cauda nuclei caudati(尾状核尾)Tail of caudate nucleus
→尾状核尾は視床と分界条によって境される。側脳室の中心部の底面の外側縁を後走し、ついで側脳室の弯曲に従って前下方に開いた弓を描きつつ下角の上壁に至り、扁桃体の後端部の外側部に接して終わる。
14. Gyrus temporalis medius(中側頭回)Middle temporal gyrus
→側頭葉には大脳回が3列に並んでいる。つまり中央の上側頭溝と下側頭溝に挟まれた脳回が中側頭回である。
15. Sulcus parieto-occipitalis(頭頂後頭溝)Parieto-occipital sulcus
→楔前部の前部は、上方に屈曲した帯状回の後部によって境され、後部は頭頂後頭溝によって境されている。
16. Sulcus calcarinus(鳥距溝)Calcarine sulcus
→一次感覚領において、楔部下方にある深い溝。楔部の下前方で頭頂後頭溝と合す。
17. Gyri occipitales(後頭葉の大脳回)Occipital gyri
→後頭葉の大脳回を一括して後頭葉の大脳回と呼んでいる。
18. Polus occipitalis(後頭極)Occipital pole
→後頭葉の後端
19. Crus anterius capsulae internae(内包前脚)Anterior limb of internal capsule
→内包前脚はレンズ核と尾状核頭部との間にある。内包前脚には、前視床縫線、前頭前皮質橋核路が含まれる。
20. Fornix(脳弓)Fornix
→白い線維が帯状になった脳弓は海馬体の主要な遠心性線維系を構成する。この中には投射線維と交連線維の両者が含まれる。これは海馬台皮質(海馬台前部、海馬台、前海馬台)と海馬の大錐体細胞の軸索からなり、海馬白質として側脳室表面に広がり、それらがまとまって海馬采を形成する。両側の海馬采は後方へ進むにつれて太くなり、海馬の後端に至って脳梁膨大の下を脳弓脚となって子を描いて上がると同時に両側の物が互いに近付いてくる。このあたりで多数の線維が反対側の脳弓に入る。すなわち交叉線維が薄く板状に広がって脳弓交連(海馬交連、または脳琴psalterium)を形成するがヒトでは発達が悪い。両側の脚は合して脳弓体となり脳梁の直下を前方に視床の吻側端まで行き、ここで再び線維束が左右に分かれ脳弓の前柱として室間孔から前交連の後ろまで腹方に曲がる。神経線維が薄い帯状になった海馬采は脳弓のほぼ全経過にわたって外側に位置しているが、吻側では脳弓の本体である脳弓前柱の中に混ざってしまう。脳弓線維の最大部分は前交連の尾側を交連後脳弓として下行し、残りは前交連の前を交連前脳弓となって走る。海馬台に起始をもつ交連後脳弓線維は視床下部を通過して乳頭体に至るが途中視床にも枝をだす。乳頭体では主として内側核に終止する。視床下部の吻側部で交連後脳弓から分かれた線維は、外側中隔核と、視床の前核群や外側背側核に終わる。その他、海馬台からの遠心線維は、前頭葉内側皮質、尾側帯状回、海馬傍回などに直接投射する。交連後脳弓線維の一部は乳頭体を越えてさらに尾方へ下り中脳被蓋に入る。交連前脳弓は線維束としては交連後脳弓よりも小さく、疎であって肉眼的に認められるようなものではない。これらの線維は海馬のすべてのセクターの錐体細胞に始まり、中隔核群の尾部の一部に終わる。以上にのべた解剖学的な連絡様式から示唆されるように、海馬台からのインパルスは複雑な経路を経て種々の部位に投射される。すなわち、海馬と海馬台は直接あるいは関節の投射路によって中隔核群、視床下部、視床、大脳皮質の広汎な領域および中脳網様体と結合する。
21. Genu capsulae internae(内包膝)Genu of internal capsule
→内包膝は前および後脚の間にある。皮質延髄路および皮質網様体路の線維が含まれる。
22. Globus pallidus(淡蒼球)Globus pallidus
→淡蒼球はレンズ核の最内側部を占め、被殻よりは小さい。垂直に走る板状の有髄神経線維(外側髄板)によって外側部の被殻と隔てられており、また、内側髄板によって内節と外節に分かれる。系統発生学的に線条体よりは古く、下等動物でよく発達している。発生に関しては、「間脳」性とするものののほか、その一部を「終脳」由来とするものがある。鉄反応が強陽性にでることが知られている。淡蒼球には大型の紡錘形ニューロンが多く、樹状突起は無棘で、長く髄板に平行して円板状に分枝する。淡蒼球の大型ニューロンの定量分析によれば、これらの細胞は単一のニューロン群に属する。淡蒼球の内節と外節の大型ニューロンには、なんら形態学的又は化学的な差異は見られない。ヒトでは外節は淡蒼球全体の約70%を占め、細胞密度が最も高い。淡蒼球の内節、外節の大型ニューロンはすべてGABA作働性である。淡蒼球ニューロンの軸索は少数の側枝を出している。淡蒼球の内側・外側髄板に存在する大型のコリン作働性ニューロンは、淡蒼球の腹側に存在する無名質のつづきである。多数の有髄線維束が淡蒼球を横走しているので、新鮮な脳では被殻や尾状核に比べ色が淡く見える。淡蒼球への求心性神経線維のもっとも主な起始は尾状核と被殻である。また、視床下核や黒質からの神経線維が内包を横切って主として内節に入る。淡蒼球が大脳皮質からの求心線維を受けるかどうかについては不確かである。視床からの求心線維については否定的な見解が多い。淡蒼球からおこる遠心性神経線維の主なものは視床と視床下核におわる。これらのうち、内節の腹側から出る神経線維は淡蒼球の腹側表面に集合してレンズ核ワナを形成し、内方後脚の腹内側部を背方にまわり、背尾側方へ走ってフォレル野に達する。一方、内節の背側からでる神経線維は淡蒼球の背内側部からおこる。これらは多数の小線維束として内包の腹側部を横切り、不確帯の腹側部に集合してレンズ核束(H2)を形成する。ついで、レンズ核束はフォレル野に入り、ここでレンズ核ワナの神経線維と一つになり、不確帯の内側端を取り囲むように腹側から背側にまわり、視床束(H1)を形成する。視床束は不確帯の背側を吻外方へ走り、視床に入って主として前腹側核(VA)、外腹側核(VL)、正中中心核(CM)などに分布する。なお、視床束には小脳視床線維も含まれている。淡蒼球から視床下核にいたる神経線維は主として外節からおこり、内包の尾側レベルにおいてその腹内側部を横切って視床下核に達する。淡蒼球からおこり中脳被蓋(脚橋被蓋核)や黒質へ向かう遠心性神経線維もある。
23. Vermis(小脳虫部)Vermis
→小脳のなかで系統発生学的に古い無対の部分。
24. Area striata(視覚皮質[有線野])Visual cortex
→視覚皮質は最も薄く、一般に顆粒細胞に冨み、特に第4層の顆粒皮質の発育が良い。また外Baillarger線条が厚く明瞭で、特にGennariまたはVicq d'Azyr線条とよばれ、肉眼的にも白い線条にとして認められる。Brodmannの17野に相当する。
25. Caput nuclei caudati(尾状核頭)Head of caudate nucleus
→尾状核頭は側脳室の前角の中に膨隆し、その外側壁をなし、視床の前方に位置する。尾状核頭は後方にしだいに小さくなり、視床の出現とともにその背外側に位置するようになり、尾状核尾に移行する。
26. Foramen interventriculare(室間孔、モンロー孔)Interventricular foramen
→『モンロー孔』ともよばれる。左右の側脳室と第三脳室とを結ぶ連絡孔。スコットランドの医学者Alexander Monro II (1733-1817)により、1797年に発見された。彼の名前はモンロー・リヒター線Monro-Richter lineにも残っている。
27. Nuclei anteriores thalami(視床前核)Anterior nuclei of thalamus
→前核群は視床の最も吻側で2分した内髄板によって囲まれた領域で背側表面の下にあり、ここに前結節を形成している。大きな主核である腹側前核(AV)と副核である背側前核(AD)および内側前核(AM)の3つの神経核が含まれる。ヒトでは腹側前核以外は退化的である。これらの諸核を構成している細胞は、中等大で、色素親和性の物質はほとんどなく、中等量の黄色色素が含まれ、有髄神経線維にとり囲まれている。乳頭体から線維を受け、帯状回と相互に線維連絡する。大脳辺縁系と関連しており、情動や新しい記憶と関連するという。
28. Nuclei mediales thalami(視床内側核)Medial nuclei of thalamus
→視床内側核は視床内髄板と髄板内核に取り囲まれている。その内方および腹内方には視床正中線核群(midline thalamic nuclei)が分布する。正中線核群は菱形核、結合核、および紐傍核である。視床内側核は内側の巨大細胞部(magnocellular part)と外側の小細胞部(paraveocellular part)に区分される。小細胞部と内髄板の境界部に大型細胞から成る髄板傍部(pars paralemellaris)が記載されることがあるが、これはむしろ視床外側中心核(CL)に属するものと考えられる。視床内側核は腹側線条体からの投射もうける。この経路によって「側坐核(腹側線条体のもっとも主要な部分)-視床内側核-前頭前野皮質」の連絡系が成立する。視床から腹側線条体への投射線維は紐傍核(parataenial nucleus)から起こる。
29. Sulcus centralis(中心溝)Central sulcus
→『ローランド溝』ともよばれる。中心溝は大脳半球の上縁から外側溝の方向に向かって下前方に走る明瞭な溝である。通常、この溝は2か所で屈曲し、上方では大脳半球の内側面までは伸びていない。この溝の深部が前頭葉と頭頂葉の境となっている。一般に、中心溝は外側溝(シルビウス裂)に達しないことが多く、内側面に入るとすぐに終わることで確認される。イタリアの解剖学者Luigi Rolando (1773-1831)による。
30. Gyrus postcentralis(中心後回)Postcentral gyrus
→中心溝とその後方に平行に走る中心後溝があり、この両溝に挟まれた脳回を中心後回(一次体性感覚野primary somatic sensory cortex)という。領域3は中心溝の後壁に沿って位置する。中心溝の後壁をなす3bと、溝の深部にある3aが領域3を構成する。領域3と領域1,2とは皮質間結合で結ばれている。領域1,2から運動野、頭頂連合野へ投射し、逆に運動野からの投射をうける。温、痛覚や触覚などの体性感覚の中枢で、内側毛帯、脊髄視床路、三叉神経毛帯を経過して、視床の後外側腹側核、後内側腹側核で中継された上行性投射線維をうける。一次体性感覚野のすべての視床投射は体性感覚局在的に構成されている。
31. Nucleus lateralis posterior(視床後外側核)Lateral posterior nucleus of thalamus
→視床の他の核から線維を受け、上・下頭頂小葉(頭頂連合野)に線維を送っている。これにより、感覚情報は連合野で解析・統合される。
32. Sulcus circularis insulae(島輪状溝)Circular sulcus of insula
→島の境界となる溝。島限で中断する。
33. Pulbinar thalami(視床枕核)Nuclei pulvinares of thalamus
→視床枕核は霊長類ではよく発達しており、前核、内側核、外側核、および下核を区別できる。視床の後部と背外側部を形成する大きな灰白質塊で、これの尾方は、内側膝状体、外側膝状体および中脳の背外側面の上に張り出している。視床枕核は、細胞学的にはかなり均一であるので、局所的な位置関係を基にして細分される。他の視床核から線維を受け、頭頂葉や後頭葉に投射する。視覚・聴覚・立体の認知にあずかるとされている。
34. Nuclei habenulae (手網核)Habenular nuclei
→嗅脳の反射路に関与する細胞群。
34a. Nucleus habenularis lateralis(外側手綱核)Lateral habenular nucleus
→外側手綱核は、淡蒼球、視床下部外側や、無名質および外側視索前野から、ならびに腹側被蓋野と中脳の縫線核からの求心性線維を受ける。
34b. Nucleus habenularis medialis(内側手綱核)Medial habenular nucleus
→手綱の小さい方の内側手綱核は、中隔核の後部と中脳の縫線核からの求心性線維を受ける。縫線核からのセロトニン作働性の投射と上頚神経節からのアドレナリン作働性支配は手綱の手綱内層核に達する。外側手綱核と内側手綱核は、中脳の吻側部へのインパルスを伝える大脳辺縁系の伝導路が集まるところである。
35. Hippocampus(海馬)Hippocampus
→海馬は(海馬はウマの前半身と長い魚の尾をもったギリシャ神話における動物)またはアンモン角(アンモンはエジプトの巻いた角をもった神)は海馬溝による長い弯曲したたかまりで、側脳室下角の内側壁の大部分を占め、その肥厚した前端部の上面にはいくつかの指のような肥厚があり、これは海馬足または海馬指とよばれ、ほとんど下角の前端に達している。海馬の側脳室表面は海馬白板によっておおわれる。これは内側方は海馬采に続く。海馬采は海馬をおおってその内側部に付く。大脳半球内側面の部分から形成される古い皮質(原皮質)で解剖学的に海馬を厳密に定義することはむずかしい。すでに爬虫類で海馬に相当する皮質がみられ、哺乳類でみられる基本的な要素が発達している。ある種の出力と入力が共通しており、互いに密接に関連(結合)している単純な層形成をもっている部分、すなわち、固有の海馬(アンモン角)と歯状回および、ときに海馬台(海馬支脚、海馬床)、海馬采、脳弓を加えて考えた方が研究目的上都合がよいことが多い。また、しばしばこの意味で、海馬形成(hippocampal formation)という言葉が用いられる。さらに皮質分野27(海馬支脚前野),29e(脳梁膨大後野)、49(海馬支脚傍野),28(内嗅領),からなるretrohippocampal foramtionを含めて海馬領域(Angevin, 1965)とよばれる。海馬は構造上部位差がみられ、CA1~CA4の亜核に分けられる(Lorente de No, 1933, 1934)。また、構造上、分子層、錐体細胞層および多形細胞層の各層に区別される。海馬からの遠心性線維は、主として前部から中核部に、また、主として後部から脳弓線維となって視床下部とくに乳頭体に終わる。求心性線維には、嗅内野(entorhinal area, 28野)や嗅周野からくる穿通線維および中隔核や対角帯核からくる中隔海馬線維がある。嗅脳からの直接投射はない。海馬は以前、嗅覚系に関係すると考えられ、ついで情動に、最近では臨床的知見により記憶、とくに短期記憶に関係があることが示唆している。側頭葉基底部の皮質および海馬を両側性に除去すると最近のできごとに関係する記憶が失われる。患者はまったく正常に会話についていけるが、話題が変わるとたちまち前の話の筋を忘れてしまう。
36. Fimbria hippocampi(海馬采)Fimbria of hippocampus かいばさい
→海馬采は脳弓脚に続いて前下方に海馬の内側部と癒着しつつ走って海馬傍回の鈎に至る。脳弓は主として海馬、海馬台などから出て海馬白板を通って脳弓に入り、乳頭体などに至る海馬乳頭路からなる。
37. Gyri temporales transversi(横側頭回、ヘッシェル横回)Transverse temporal gyri
→ヘッシェル横回とも呼ばれる。上側頭回の背側面にあって外側溝の底にかくれて横側頭回のがある。横側頭回の内側部には皮質聴覚中枢(聴覚野)があり、その外側部から上側頭回の表面にかけて聴覚性連合野が広がる。Heschl, Richard (1854-1881) オーストリアの病理学者。グラーツ、ウェーン大学の教授を歴任。ヘッシュル横回(横側頭回)を記述、グラーツ時代に病理標本館を作る。論書は50篇以上になるが、特異なものにロキタンスキーの行った病理解剖法を著したものがある("Sectionstechnik", 1859)。
最終更新日: 19/02/05