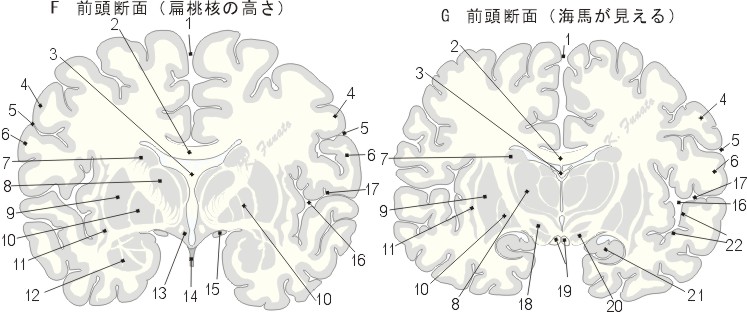
この高さでは背尾側から腹吻側へ斜めに走っている中心溝の切れ目がみえる。この中心溝は比較的遠く離れた吻側で切断されるため、その背側にある前頭葉はその腹側にある頭頂葉よりも、この断面では著しく広い部分を占めることになる。中心溝の上にある回は中心前回であって、その下にある回は中心後回である。側頭葉の深部には扁桃体(扁桃核複合)、が現れる。扁桃体は側頭葉の内側面で表面にまで達しているため、これは一部は皮質として、また一部は核とみなされることもあるが、両構造の移行型とみるのがより適切であろう。まわりの扁桃周囲野のみならず、扁桃体の皮質内側核群も第1次嗅覚中枢に属しているから、その核としての性格をもつにかかわらず、扁桃体を古皮質に算入するのである。扁桃体の上には広い底をなして前障が終わっている。両半球の間には今や視床、淡蒼球および視床下部からなる間脳がみえてくる。これら間脳の諸核の外側には、被殻と尾状核を合わせた線条体が接している。脳梁の下には、脳弓の太い線維束がみられる。その他、大脳縦裂、大脳外側溝、外側窩、視索および漏斗も見える。
扁桃体の高さ(前頭断面) Fに対する面
01:大脳縦裂、02:脳梁、03:脳弓、04:中心前回、05:中心溝、06:中心後回、07:尾状核、08:視床、09:被殻、10:淡蒼球、11:前障、12:扁桃体[扁桃核複合]、13:視床下部、14:漏斗、15:視索、16:外側窩、17:大脳外側溝、18:視床下核、19:乳頭体、20:黒質、21:海馬、22:横側頭回
01. Fissura longitudinalis cerebri(大脳縦裂)Longitudinal cerebral fissure
→左脳半球は右大大脳縦裂によって不完全に分けられており、生体ではこの部分に大脳鎌が入り込んでいる。前頭部と後頭部では大脳半球は完全に分かれているが、中央部では大脳縦裂が幅広い半球間の交連線維群である脳梁までしか達していない。
02. Corpus callosum(脳梁)Corpus callosum
→脳梁は左右の大脳皮質、ことに新皮質を結合する線維の集合したもので、系統発生的には最も新しく、ヒトでは非常に発育がよい。その前後経はほぼ7.7cmである。脳梁は正中断では全体としては釣針状で、4つの部分が区別される。後端部は膨大し、脳梁膨大といい、その前方に続いて水平に走る部分を脳梁幹とよぶ脳梁はその前端では強く屈曲し、脳梁膝をつくる。これはさらに後下方にくちばしのように尖って脳梁吻となり、しだいに薄くなって終板に続く。
03. Fornix(脳弓)Fornix
→白い線維が帯状になった脳弓は海馬体の主要な遠心性線維系を構成する。この中には投射線維と交連線維の両者が含まれる。これは海馬台皮質(海馬台前部、海馬台、前海馬台)と海馬の大錐体細胞の軸索からなり、海馬白質として側脳室表面に広がり、それらがまとまって海馬采を形成する。両側の海馬采は後方へ進むにつれて太くなり、海馬の後端に至って脳梁膨大の下を脳弓脚となって子を描いて上がると同時に両側の物が互いに近付いてくる。このあたりで多数の線維が反対側の脳弓に入る。すなわち交叉線維が薄く板状に広がって脳弓交連(海馬交連、または脳琴psalterium)を形成するがヒトでは発達が悪い。両側の脚は合して脳弓体となり脳梁の直下を前方に視床の吻側端まで行き、ここで再び線維束が左右に分かれ脳弓の前柱として室間孔から前交連の後ろまで腹方に曲がる。神経線維が薄い帯状になった海馬采は脳弓のほぼ全経過にわたって外側に位置しているが、吻側では脳弓の本体である脳弓前柱の中に混ざってしまう。脳弓線維の最大部分は前交連の尾側を交連後脳弓として下行し、残りは前交連の前を交連前脳弓となって走る。乳頭体では主として内側核に終止する。視床下部の吻側部で交連後脳弓から分かれた線維は、外側中隔核と、視床の前核群や外側背側核に終わる。その他、海馬台からの遠心線維は、前頭葉内側皮質、尾側帯状回、海馬傍回などに直接投射する。交連後脳弓線維の一部は乳頭体を越えてさらに尾方へ下り中脳被蓋に入る。
04. Gyrus precentralis(中心前回)Precentral gyrus
→大脳半球の外側面で、中心溝のすぐ前にある高まりが中心前回で、ここは随意運動に関係する運動領(運動野)motor areaである。
05. Sulcus centralis(中心溝)Central sulcus
→『ローランド溝』ともよばれる。中心溝は大脳半球の上縁から外側溝の方向に向かって下前方に走る明瞭な溝である。通常、この溝は2か所で屈曲し、上方では大脳半球の内側面までは伸びていない。この溝の深部が前頭葉と頭頂葉の境となっている。一般に、中心溝は外側溝(シルビウス裂)に達しないことが多く、内側面に入るとすぐに終わることで確認される。イタリアの解剖学者Luigi Rolando (1773-1831)による。
06. Gyrus postcentralis(中心後回)Postcentral gyrus
→中心溝とその後方に平行に走る中心後溝があり、この両溝に挟まれた脳回を中心後回(一次体性感覚野primary somatic sensory cortex)という。領域3は中心溝の後壁に沿って位置する。中心溝の後壁をなす3bと、溝の深部にある3aが領域3を構成する。領域3と領域1,2とは皮質間結合で結ばれている。領域1,2から運動野、頭頂連合野へ投射し、逆に運動野からの投射をうける。温、痛覚や触覚などの体性感覚の中枢で、内側毛帯、脊髄視床路、三叉神経毛帯を経過して、視床の後外側腹側核、後内側腹側核で中継された上行性投射線維をうける。一次体性感覚野のすべての視床投射は体性感覚局在的に構成されている。
07. Nucleus caudatus(尾状核)Caudate nucleus
→尾状核は全体として弓状の大きな灰白質である。の吻側部は視床の吻側に位置し、側脳室前角のなかに隆起してその外側壁をなし、尾状核頭とよばれる。後方で細くなって尾状核尾とよばれ、視床の背外側縁に沿って側脳室の中心部の底面の外側縁を走り、やがて側脳室の弯曲に沿って下方にまがり、側脳室下核の上壁に達し、扁桃体との後端部のレベルでその外側部に接しておわる。尾状核頭と尾状核尾の中間部を尾状核体という。神経細胞には、大きく分けて、大小2種類(1:20)のものがみられる。求心性神経線維の起始部の主なものは、大脳皮質・視床髄板内核・視床正中中心核・黒質である。黒質(とくに緻密部)からはドパミン作働性の神経線維を受ける。尾状核からの遠心性神経線維の主な分布域は淡蒼球と黒質(とくに網様部)であり、これらの多くのものはGABA作働性である。
08. Thalamus(視床)Thalamus
→視床は、背側間脳溝と視床下溝の間の領域であるが、発生の間に大きく発育して、間脳背側部の広い範囲を占めるようになる。間脳は個体発生上、背側視床、腹側視床、視床下部および視床上部の四つの部位に分けられるが、その中で最も大きな部位を占めるのが背側視床である。単に視床といった場合は背側視床を指す。視床は第三脳室の両壁をなす卵円形の構造で、背側の遊離面は薄い髄質から成る帯層におおわれ、肺内側端に視床上部の構造である視床髄条が、前端より後方に走り手網核に付く。また背外側端は分界条によって終脳の尾状核と、外側方は外髄板によっておおわれ腹側視床の視床網様核と境されている。左右の視床は第三脳室内にまたがる視床間橋(中間質)によってつながり、視床下溝で視床下部と境される。視床の内部を構成している視床核は視床脚を介して大脳皮質と相互に結合する。内部には内髄板とよばれる線維板視床を内側部、外側部および前部に分けている。視床は感覚系と統合系との非常に重要な連絡部位である。嗅覚路以外のすべての感覚路がそれぞれ相当する視床の領域に投射する。「最近の研究によれば、嗅覚系も視床を投射する可能性がある」。視床で処理された感覚系情報の流れは視床大脳皮質線維を経て大脳皮質へと送られるが、大脳皮質の側からは多数の大脳皮質視床線維を介して視床における情報処理系に影響が及んでおり、したがって、視床と大脳皮質とは一つの機能単位としてはたらく。「運動」情報は小脳と大脳基底核を経て伝達され、統合系(大脳辺縁系や脳幹網様体など)からのさらに複雑な情報も視床に達する。したがって、視床は一方では大脳辺縁系と脳幹網様体との連結点として機能し、他方では大脳皮質も連絡しているわけである。
09. Putamen(被殻)Putamen
→→レンズ核の外側部。終脳から発生した部分。被殻はレンズ核の外側部を形成し、外側髄板によって淡蒼球の外節とへだてられている。島皮質とは最外包、前障、外包によってわけられる。被殻の構造は尾状核とまったく同様で、太い有髄線維をほとんど含まず、主として小さい神経細胞からなるが、散在性の大細胞を含む。被殻と尾状核は発生学的にみると、同一の細胞群が内包の発達によって隔てられたもので、両者の間には互いに結合する灰白質の線条が多数見られる。そのため、両者をあわせて線条体または新線条体と呼ぶ。線維連絡も尾状核と原則的に等しい。霊長類において動物が高等になると、相対的な意味で尾状核の体積が減少し、被殻の体積が増大するといわれている。
10. Pallidum(淡蒼球、古線条体、旧線条体)Pallidum; Paleostriatum
→淡蒼球は薄い内側髄板によってさらに内節と外節に分けられるが、両部は構造上の差異はなく、いずれも散在性の大きい細胞からなり、有髄線維に富み、肉眼的にやや白く見える。
11. Claustrum(前障)Claustrum
→前障はレンズ核と島との間にある、内側が凹面をなす板状の核で、腹側方に厚くなる。この核とレンズ核との間には外包があり、また島の皮質との間には最外包がある。これらは狭い白質で、大部分は連合線維から、一部は交連および投射線維からなる。前障は種々の視床核、扁桃体などから線維を受け、大脳皮質に広く投射する。前障は以前は線条体とともにいわゆる基底核に数えられたり、あるいは皮質層の付け足しとして島皮質に属するものとされた。しかしながら、発生学的ならびに比較解剖学的研究によって、前障は発生の途中で位置がずれた古皮質の細胞群であることが証明されている。前障はその広い底の所で古皮質の領域へ移行する(すなわち梨状前野や扁桃体の外側核へ)。頭頂葉、側頭葉および後頭葉の皮質からの、無髄線維が局在的配列をなして前障に終わると言われている。前障の機能についてはわかっていない。
12. Corpus amygdaloideum(扁桃体、扁桃核、扁桃体複合核)Amygdaloid body; Amygdaloid complex
→扁桃体または扁桃核はアーモンドに似た形をした系統発生学的に古い核で、原始線条体とも呼ばれ、辺縁系における重要な神経核をなしている。扁桃体は側脳室下角の前端部の前にあり、一部その前内側壁と前背側壁を作り、前頭断では大きい不規則な卵円形の細胞集団をなし、レンズ核の腹側にある。この核は多数の亜核からなり、系統発生学的に古い内側核群とやや新しい外側核群に分かたれる。扁桃核は周辺の皮質や嗅球、その他の嗅脳部、中隔野などから線維を受けるが、海馬からの直接の線維はないらしい。扁桃核の遠心路には背側の分界条と腹側の遠心系がある。分界条は扁桃核から起こって尾状核尾の内側縁に沿って上方、ついで前方に向かい、視索前部、視床下部の前核および腹内側核に分布する。腹側遠心系はレンズ核の腹側を内側方、前方に走り、視索前部、中隔野、尾状核頭腹側部、嗅脳後部、前交連などになどに入り、一部の線維は内側前脳束に入って主として視床下部外側核に分布し、また一部は下視床脚を通って視床内側核に入る。扁桃体は視床・視床下部や前頭葉・側頭葉の新皮質との間に複雑な線維連絡をもつ。扁桃体は本能・情動による行動の中枢で、その線維連絡によりいろいろな体性および内臓機能と関連すると考えられている。
13. Hypothalamus(視床下部)Hypothalamus
→第三脳室の側壁の下部および底にあたる。脳底面からみると、吻側から数えて、視交叉、漏斗、灰白隆起、乳頭体とつづき、漏斗の先端は下垂体に連なる。背側は視床下溝により視床と境されており、吻側は終脳の視索前野に、尾側は中脳被蓋と中脳中心灰白質に、尾外側は視床腹側部に移行する。通常、矢状面に平行な三つの帯状領域、すなわち、視床下部脳室周囲層、視床下部内側野、視床下部外側野に区分される。これらの間を多数の細かい神経線維が主として吻尾方向に走っている。視床下部は前後径が約10mmである。視床下部は内部環境を正常に維持する機序に関与し、また心悸亢進、瞳孔散大、“冷汗”の分泌などの情動反応の表出にも一役を演じている。大脳皮質を除去し、背側視床を除去した後でも、怒り反応はあらわれる(「みかけの怒りSham rage」)。さらに、視床下部は成長、性的成熟など新地あの成熟過程にも関与している。したがって、視床下部を損傷すると、広汎で顕著な内分泌性、代謝性、行動性(情動性)の以上が一緒に起こってくることになる。視床下部は下垂体ホルモンによって内部環境に影響を及ぼすが、さらに脳幹網様体や自律神経系を介しても同様の働きを示す。視床下部への情報は通常の入力神経系によって伝達されるばかりでなく、視床下部のニューロンは内部環境からの物理的および化学的刺激(血液の温度など)にも反応する。植物性機能の中には、その統御中枢が視床下部自体に存在するものもあるが、呼吸や心臓の活動などの機能に関しては、視床下部はもっと回の中枢に対する修飾器として働いている。
14. Infundibulum(漏斗)Infundibulum
→視床下部の腹側方の突出部とその中にある第三脳室の陥凹によって、漏斗が形成される。漏斗の最も遠位に突出した部分が下垂体後葉(神経下垂体)であり、漏斗の突出部と正中隆起を結合する組織は漏斗柄とよばれる。
15. Tractus opticus(視索)Optic tract
→左右の視索は視床下部と大脳脚の吻側部一部を巡る。これらの線維の多くは外側膝状体の中に終止するが、小部分は下丘腕となって上丘および視蓋前域にまで続く。外側膝状体からは膝状体鳥距路が起こり、これが視覚路の最後の中継路をなす。視索前域は対光反射と関係し、上丘は眼と頭の反射運動よよち視覚刺激を追跡することと関係している。網膜視床下部線維は、両側性に視床下部の視神経交叉上核に終止する。この網膜からの直接の投射は、機能的には神経内分泌調節と関連している。
16. Fossa lateralis cerebri(大脳外側窩)Lateral cerebral fossa
→外側溝深部にある腔所。外側溝は下面における陥凹である大脳外側窩の外側端ではじまり、しばらく上外側方に走って外側面に現れ、ここで3つの枝に分かれる。
17. Sulcus lateralis(外側溝)Lateral sulcus
→『シルビウス裂溝』ともよばれる。外側口はぢあの右半球の底面における陥凹である大脳外側窩に始まり、外包にすすんで半球外側面に現れ、その主部は後枝として後上方にすすみ、一方は前頭葉および頭頂葉と他方は側頭葉との境をなす深い溝である。半球外側面に現れたところで2小枝、すなわち前に向かう前枝と、上行する上行枝を出す。外側溝の奥には島がある。オランダの医学者Francis Sylvius (1614-1672)による。ちなみに中脳水道のシルビウスは別人である。
18. Nucleus subthalamicus(視床下核)Subthalamic nucleus
→ルイ核ともよばれている。脳の断面の肉眼観察の際にも「目立つ」神経核であって、大型ニューロンから成り、間脳の最尾部において内包後脚の背内方に位置している。この神経核の内側部は黒質吻側部の背方に位置する。背側の不確帯とはレンズ核束(H2)によりへだてられている。核の境界は明瞭で、前頭断面では両凸レンズ形を呈し、矢状断面ではほぼ円形を呈する。核の尾側端のレベルでは、核の内側部が黒質の最吻側端の背縁に接している。主な求心出力神経線維を淡蒼球や脚橋被蓋核受け、また、遠心性神経線維を主として淡蒼球内節に送る。大脳皮質とくに前頭葉からの求心性線維や、黒質や淡蒼球への遠心性線維の存在が報告されているが、その他の線維連絡関係については不確実な点が多い。ヒトでこの核が損傷されると、反対側の半身に激しい不随意運動、すなわちヘミバリスムがおこる。視床下核の細胞はグルタミン酸塩を含有しており、淡蒼球と黒質のニューロンに興奮性に作用すると言われている。グルタミン酸塩は細胞の基礎代謝にも存在するもので、視床下核の細胞がグルタミン酸塩免疫反応陽性であっても、かならずしもグルタミン酸塩がこの核の細胞により使用される神経伝達物質とはいえない。視床下核は視床下部外側核の最後の細胞集団から発生してくる。吻側の細胞集団は淡蒼球の内節、外節の原基となる。Luys, Jules Bernard (1828-1898)フランスの神経学者。
19. Corpora mamillaria(乳頭体)Mamillary body
→乳頭体は有髄線維を豊富に含み、視床下部の中では特物の位置を占めている。乳頭体は内側および外側乳頭体核より成る。内側乳頭体核は外側乳頭体核より大きいが、そのニューロンは比較的小さく、有髄線維のカプセルに包まれている。外側乳頭体核はずっと小さくて、「とくにヒトでは、見分けるのがむずかしい。」そのニューロンは内側乳頭体核のものよりも大きく、染色されやすい。乳頭体への入力線維は、海馬支脚(交連後部脳弓を介して)、視床下部腹内側核、中脳(乳頭体脚を介して)、などからくる。脳弓の線維数は非常に多い。脳弓線維は内側乳頭体核に終止するが、中には乳頭体を通り越して中脳被蓋ないし中心灰白質でシナプス結合するものもある。中脳から乳頭体への入力線維は、中脳中心灰白質および中脳網様体にある楔状核から起こり、乳頭体脚を通ってくる。乳頭体からの出力線維は大脳辺縁系の重要な要素である。内側乳頭体核から出る線維は明瞭な上行性線維束である主乳頭体束を作り、これは吻背側方へ向かう乳頭体視床路と、尾側方へ向かうこれよりも小さい乳頭体被蓋束に分かれる。乳頭体視床路線維は主として内側乳頭体核より起こり、視床前核群でシナプス結合する。視床前核は帯状回でシナプス結合する線維を出す。「海馬→脳弓→乳頭体→視床前核→帯状回」の回路は大脳辺縁系の中心的な回路としてしられている(Papezの情動回路)。乳頭体被蓋束の出力線維は内側乳頭体の背側部より起こり、中脳被蓋でシナプス結合する。
20. Substantia nigra(黒質)Substantia nigra
→中脳被蓋腹側部の核で大脳脚の背側に接して存在する。ヒトの黒質の神経細胞は顆粒状のメラニン色素を豊富に含有するため、黒質は全体として肉眼的に黒くみえる。黒質には背側の緻密部と腹側の網様部が区分される。緻密部が神経細胞に富むのに対し、網様部では神経細胞の密度は粗で、細い神経線維に富む。したがて、前者は黒色部、後者は赤色部とよばれることがある。黒質からおこる遠心性神経線維としては、緻密部からおこり線条体に分布する黒質線条体線維、網様部から起こり視床のとくに内側腹側核(VM)に分布する黒質視床線維、および網様部からおこり上丘の中間灰白質に分布する黒質上丘線維などが主なものである。また、黒質に分布する求心性神経線維の起始としては、線条体・淡蒼球・視床下核(Luys体)が主なものである。これらのほか、前頭葉皮質・背側縫線核・扁桃体中心核・外側手綱核なども報告されているが不確実である。黒質は中枢神経系のうちでドーパミンとGABAの含有量が高い部位として知られる。ドーパミンは線条体に神経線維を送る黒質緻密部の神経細胞に主として含まれ、またGABAは線条体よりおこり黒質網様体に至る神経線維の軸索終末に主として含まれる。黒質に見られる線維はまたは11個のアミノ酸が連絡したペプチドとしてのP物質(SP)も含む。黒質は脳において最も高濃度にP物質を有する部位で、この物質は黒質の緻密部および緻密部内の神経終末に凝集している。網様部はまたエンケファリン作働性線維および終末も有する。尾状核および被殻の樹状突起の棘突起に含むニューロンから起こる線条体黒質線維はGABA、P物質、エンケファリンを含む。これらの線維は同様の伝達物質を有する線条体淡蒼球線維を出すニューロンとは異なる細胞集団から起こる。黒質はパーキンソン病(振戦麻痺)の原因となっている代謝障害に緻密に関係しており、Huntington舞踏病および異常な不随意運動や筋緊張の変化を特徴とする他のタイプの運動障害にも関与しているようである。パーキンソン病では黒質から線条体へのドーパミンの輸送および合成が極度に傷害される。Huntington舞踏病では線条体のドーパミンは星状であるがGABAは著明に減少している。
21. Hippocampus(海馬)Hippocampus
→海馬は(海馬はウマの前半身と長い魚の尾をもったギリシャ神話における動物)またはアンモン角(アンモンはエジプトの巻いた角をもった神)は海馬溝による長い弯曲したたかまりで、側脳室下角の内側壁の大部分を占め、その肥厚した前端部の上面にはいくつかの指のような肥厚があり、これは海馬足または海馬指とよばれ、ほとんど下角の前端に達している。海馬の側脳室表面は海馬白板によっておおわれる。これは内側方は海馬采に続く。海馬采は海馬をおおってその内側部に付く。大脳半球内側面の部分から形成される古い皮質(原皮質)で解剖学的に海馬を厳密に定義することはむずかしい。すでに爬虫類で海馬に相当する皮質がみられ、哺乳類でみられる基本的な要素が発達している。ある種の出力と入力が共通しており、互いに密接に関連(結合)している単純な層形成をもっている部分、すなわち、固有の海馬(アンモン角)と歯状回および、ときに海馬台(海馬支脚、海馬床)、海馬采、脳弓を加えて考えた方が研究目的上都合がよいことが多い。また、しばしばこの意味で、海馬形成(hippocampal formation)という言葉が用いられる。さらに皮質分野27(海馬支脚前野),29e(脳梁膨大後野)、49(海馬支脚傍野),28(内嗅領),からなるretrohippocampal foramtionを含めて海馬領域(Angevin, 1965)とよばれる。海馬は構造上部位差がみられ、CA1~CA4の亜核に分けられる(Lorente de No, 1933, 1934)。また、構造上、分子層、錐体細胞層および多形細胞層の各層に区別される。海馬からの遠心性線維は、主として前部から中核部に、また、主として後部から脳弓線維となって視床下部とくに乳頭体に終わる。求心性線維には、嗅内野(entorhinal area, 28野)や嗅周野からくる穿通線維および中隔核や対角帯核からくる中隔海馬線維がある。嗅脳からの直接投射はない。海馬は以前、嗅覚系に関係すると考えられ、ついで情動に、最近では臨床的知見により記憶、とくに短期記憶に関係があることが示唆している。側頭葉基底部の皮質および海馬を両側性に除去すると最近のできごとに関係する記憶が失われる。患者はまったく正常に会話についていけるが、話題が変わるとたちまち前の話の筋を忘れてしまう。
22. Gyri temporales transversi(横側頭回)Transverse temporal gyri
→上側頭回の背側面にあって外側溝の底にかくれて横側頭回のがある。横側頭回の内側部には皮質聴覚中枢(聴覚野)があり、その外側部から上側頭回の表面にかけて聴覚性連合野が広がる。
最終更新日: 19/02/05