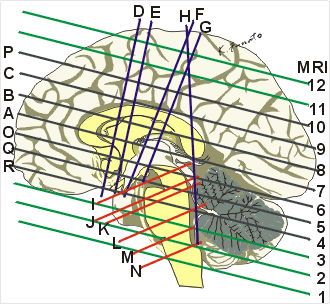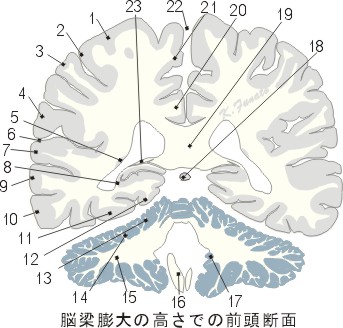
脳梁膨大部を通る面(前額断面)Hに対する面
01:中心前回、02:中心溝、03:中心後回、04:縁上回、05:尾状核尾、06:外側溝[シルビウス裂]、07:上側頭回、08:海馬、09:中側頭回、10:下側頭回、11:外側後頭側頭回、12:内側後頭側頭回、13:小脳前葉、14:第一裂、15:小脳後葉、16:菱形窩、17:第四脳室脈絡叢、18:松果体、19:脳梁膨大、20:帯状回、21:中心傍小葉、22:大脳鎌、23:脳弓
01. Gyrus precentralis(中心前回)Precentral gyrus
→大脳半球の外側面で、中心溝のすぐ前にある高まりが中心前回で、ここは随意運動に関係する運動領(運動野)motor areaである。
02. Sulcus centralis(中心溝)Central sulcus
→→『ローランド溝』ともよばれる。中心溝は大脳半球の上縁から外側溝の方向に向かって下前方に走る明瞭な溝である。通常、この溝は2か所で屈曲し、上方では大脳半球の内側面までは伸びていない。この溝の深部が前頭葉と頭頂葉の境となっている。一般に、中心溝は外側溝(シルビウス裂)に達しないことが多く、内側面に入るとすぐに終わることで確認される。イタリアの解剖学者Luigi Rolando (1773-1831)による。
03. Gyrus postcentralis(中心後回)Postcentral gyrus
→中心溝とその後方に平行に走る中心後溝があり、この両溝に挟まれた脳回を中心後回(一次体性感覚野primary somatic sensory cortex)という。領域3は中心溝の後壁に沿って位置する。中心溝の後壁をなす3bと、溝の深部にある3aが領域3を構成する。領域3と領域1,2とは皮質間結合で結ばれている。領域1,2から運動野、頭頂連合野へ投射し、逆に運動野からの投射をうける。温、痛覚や触覚などの体性感覚の中枢で、内側毛帯、脊髄視床路、三叉神経毛帯を経過して、視床の後外側腹側核、後内側腹側核で中継された上行性投射線維をうける。一次体性感覚野のすべての視床投射は体性感覚局在的に構成されている。
04. Gyrus supramarginalis(縁上回)Supramarginal gyrus
→外側溝の上行枝をとり囲む皮質部位を占め、Brodmannの領域40に相当する。縁上回は背方にある上頭頂小葉から感覚情報をうけていて頭頂連合野に属する。優位半球のこの領域が限局性に障害されると、失書、手指失認、身体部位失認が生じる。また、伝導失語や観念運動失行もしばしば認められる。
05. Cauda, nuclei caudati(尾状核尾)Tail of caudate nucleus
→尾状核尾は視床と分界条によって境される。側脳室の中心部の底面の外側縁を後走し、ついで側脳室の弯曲に従って前下方に開いた弓を描きつつ下角の上壁に至り、扁桃体の後端部の外側部に接して終わる。
06. Sulcus lateralis(外側溝)Lateral sulcus
→『シルビウス裂溝』ともよばれる。外側口はぢあの右半球の底面における陥凹である大脳外側窩に始まり、外包にすすんで半球外側面に現れ、その主部は後枝として後上方にすすみ、一方は前頭葉および頭頂葉と他方は側頭葉との境をなす深い溝である。半球外側面に現れたところで2小枝、すなわち前に向かう前枝と、上行する上行枝を出す。外側溝の奥には島がある。オランダの医学者Francis Sylvius (1614-1672)による。ちなみに中脳水道のシルビウスは別人である。
07. Gyrus temporalis superior(上側頭回)Superior temporal gyrus
→上側頭回は外側溝と上側頭溝の間にある。上側頭回の後部付近には感覚性言語中枢sensory speech center(ウェルニッケ中枢Wernicke's area)があり、これは聞いた言葉を理解する中枢であるという。一次聴覚野は聴覚の中枢で、側頭葉の上側頭回の上面(Brodmannの41野)にある。二次聴覚野は一次聴覚野の周囲(42、22野)にある。一次聴覚野で聞く音の意味はこの皮質領域で理解される。ウェルニッケ中枢の障害時には、言語の理解ができない。ちょうどしらない外国語を聞くのに似ている。また、自己の発する言語音を聞きながら発声をすることができないので、理解できるような言語を発することもできなくなる。乳児のようにチンプンカンプンの発語となる(jargon aphasia)。このように感覚性言語野の障害で起こる失語を感覚性失語症sensory aphasiaまたは言語聾word deafnessという。感覚性言語中枢と聴覚領のすぐ下の上側側頭溝には、眼で見た物の動きを総合して認識する神経細胞が存在する。一側の一次聴覚野の障害では、反対側の聴力に障害が起こるが、聴力が完全に失われることはない。一側の聴覚器からの入力は両側の聴覚野に達するためである。二次聴覚野の障害が起こると、聞く音の意味を理解することができない。これを聴覚失認auditory agnosiaという。
08. Hippocampus(海馬)Hippocampus
→海馬は(海馬はウマの前半身と長い魚の尾をもったギリシャ神話における動物)またはアンモン角(アンモンはエジプトの巻いた角をもった神)は海馬溝による長い弯曲したたかまりで、側脳室下角の内側壁の大部分を占め、その肥厚した前端部の上面にはいくつかの指のような肥厚があり、これは海馬足または海馬指とよばれ、ほとんど下角の前端に達している。海馬の側脳室表面は海馬白板によっておおわれる。これは内側方は海馬采に続く。海馬采は海馬をおおってその内側部に付く。大脳半球内側面の部分から形成される古い皮質(原皮質)で解剖学的に海馬を厳密に定義することはむずかしい。すでに爬虫類で海馬に相当する皮質がみられ、哺乳類でみられる基本的な要素が発達している。ある種の出力と入力が共通しており、互いに密接に関連(結合)している単純な層形成をもっている部分、すなわち、固有の海馬(アンモン角)と歯状回および、ときに海馬台(海馬支脚、海馬床)、海馬采、脳弓を加えて考えた方が研究目的上都合がよいことが多い。また、しばしばこの意味で、海馬形成(hippocampal formation)という言葉が用いられる。さらに皮質分野27(海馬支脚前野),29e(脳梁膨大後野)、49(海馬支脚傍野),28(内嗅領),からなるretrohippocampal foramtionを含めて海馬領域(Angevin, 1965)とよばれる。海馬は構造上部位差がみられ、CA1~CA4の亜核に分けられる(Lorente de No, 1933, 1934)。また、構造上、分子層、錐体細胞層および多形細胞層の各層に区別される。海馬からの遠心性線維は、主として前部から中核部に、また、主として後部から脳弓線維となって視床下部とくに乳頭体に終わる。求心性線維には、嗅内野(entorhinal area, 28野)や嗅周野からくる穿通線維および中隔核や対角帯核からくる中隔海馬線維がある。嗅脳からの直接投射はない。海馬は以前、嗅覚系に関係すると考えられ、ついで情動に、最近では臨床的知見により記憶、とくに短期記憶に関係があることが示唆している。側頭葉基底部の皮質および海馬を両側性に除去すると最近のできごとに関係する記憶が失われる。患者はまったく正常に会話についていけるが、話題が変わるとたちまち前の話の筋を忘れてしまう。
09. Gyrus temporalis medius(中側頭回)Middle temporal gyrus
→側頭葉には大脳回が3列に並んでいる。つまり中央の上側頭溝と下側頭溝に挟まれた脳回が中側頭回である。
10. Gyrus temporalis inferior(下側頭回)Inferior temporal gyrus
→下側頭溝の下方にある。
11. Gyrus occipitotemporalis lateralis(外側後頭側頭回)Lateral occipitotemporal gyrus
→後頭側頭溝の外側にある回。側頭葉下縁で切れ目なく下側頭回へ移行する。
12. Gyrus occipitotemporalis medialis(内側後頭側頭回)Medial occipitotemporal gyrus
→内側後頭側頭回は後頭極から側頭極まで延びている。この脳回の内側縁は側副溝と嗅溝によって境され、外側縁は後頭側頭溝によって境されている。
13. Lobus cranialis anterior cerebelli(小脳前葉)Anterior lobe of cerebellum
→小脳第一裂より前方の部分。ヒトでは古小脳症候群が確定していないが、イヌの小脳小葉を傷害すると姿勢の悪い障害と伸筋緊張の非常な増強を起こす。これらの場合、除脳に伴って見られる多くの特徴を示すが、時には歩行が可能になったり、随意運動を行うことがある。Sherringtonは小脳前葉を電気的に刺激すると除脳動物の伸筋の緊張が抑制されるという古典的実験を行った。その後の研究で小脳前葉刺激の与え方により筋緊張に対して抑制的または促進的効果が及ぼされることが明らかにされた。すなわち、低頻度刺激(2~10Hz)では同側の伸筋の緊張が緩やかに増加し、高頻度刺激(30~300Hz)では筋の弛緩が起こった。前葉刺激による抑制効果は位置小脳皮質より前庭神経外側核に至る小脳前庭繊維、②室頂核より網様体への遠心性投射により伝えられ、また促進効果は室頂核より前庭神経外側核への投射により伝えられると思われる。なおその他に、筋緊張に作用する樟脳の機序として、傍虫部皮質、中位核(栓状核)および対側の赤核が関与するものがある。これらはすべて体部位的局在をもち、相互に連絡する。ネコで中位核の前部を刺激すると、同側後肢を屈曲し、同じ核の後部を刺激すると同側前枝が屈曲した。これらの反応は上小脳脚および赤核脊髄路の線維が交叉するので、同側性に現れる。中位核より対側の赤核への投射は、赤核の細胞を興奮させ、これは赤核脊髄路を通って屈筋の筋緊張を高めるように作用する。
14. Fissura prima(第一裂)Primary fissure
→四角小葉と単小葉間の切れ目。
15. Lobus caudalis posterior cerebelli(小脳後葉)Posterior lobe of cerebellum
→第一裂と後外側裂の間になる区域。
16. Fossa rhomboidea(菱形窩)Rhomboid fossa
→菱形窩は菱形をなし、正中溝により左右に分けられ、さらにその外側の境界溝により内外の領域に分けられる。正中口からは外側に向かって第四脳室髄条が走り、これによって菱形窩はさらに上下2部に分けられる。上部では境界溝の内外に、内側隆起と前庭神経野がある。前者の中央には顔面神経丘とよぶ隆まりがあり、そこには顔面神経膝とその腹側にある外転神経核とが存在する。後者は前庭神経核の場所に相当する。前庭神経野の外側にある凹みが上窩で、これより吻側に青斑が帯状をなして伸びている。その内部に青斑核がある。菱形窩の下部の内側は舌下神経核がある。その外側には迷走神経背側核のある迷走神経三角(灰白翼)がある。その吻側端野窩みは下窩とよばれる。
17. Plexus chorioideus ventriculi quarti(第四脳室脈絡叢)Choroid plexus of fourth ventricle
→上衣組織に被われた一対の花飾り状の血管絨毛塊。外側溝まで達す。
18. Glandula pinealis; Corpus pineale(松果体)Pineal gland
→蓋板上方にあり、機能的関連のない手網についている。松果体は円錐形の小体で、後交連の領域で、第三脳室の天井に付着している。これは痕跡的な腺であるらしく、血管の豊富な結合組織の小柱の網工からなり、その網眼の中には神経膠細胞および松果体細胞pinealocytesあるいはepiphysial cellがみられる。サルの松果体細胞はセロトニン(5-HT)とコレシストキニン(CCK)を含んでいる。哺乳類の松果体細胞は、系統発生学的には、感覚神経性光受容体の要素と関係しており、これは、分泌細胞になる物が多いが、間接的に光受容性をもっている。これらの突起の棍棒状をした終末は、血管血管を取り囲む血管周囲腔に接して終わる。松果体の分泌のうちで最もよく知られた物は、セロトニン、ノルアドレナリンおよびメラトニンという生体産生アミンであるが、その他に、サイロトロピン遊離ホルモン(TRH)、黄体形成ホルモン遊離ホルモン(LHRH)およびソマトスタチン(SRIF)のような、視床下部で形成されると同定されたペプチドを、かなりの濃度で含んでいる。セロトニンは、松果体細胞の中で合成されて細胞外の隙間に放出される。ノルアドレナリンは、松果体の実質細胞に終止ししている交感神経ニューロンの中で合成される。松果体は、昼間光の変動に敏感なN-アセチルトランスフェラーゼおよびヒドロキシインドール-o-メチルトランスフェラーゼという2つの酵素の働きによって、セロトニンからメラトニンを合成する。メラトニン合成の毎日の変動は周期性であり、光入力の毎日の周期に直接関係している。光は、日周期を環境の光周期に一致させる。そして、さらに、まだ確認されていない経路を経て、神経性信号が松果体に運ばれることを速やかに遮断するように働く。N-アセチルトランスフェラーゼの活性は、昼夜に高められるが、光にさらすと、酵素の活性が失われる。視床下部の視交叉上核を両側性に傷害すると、この核は網膜視床下部経路を受けているので、松果体のN-アセチルトランスフェラーゼにおける周期がなくなり、その結果、ヒドロキシインドール-o-メチルトランスフェラーゼの活性のレベルが下がる。そのような傷害によって、自発運動の活性の日周期および栄養補給と水飲み行動の両方の日周期がなくなる。雄のネズミでは、視交叉上核の傷害によって、正常な発情周期がなくなる。網膜視床下部路は、視交叉上核の構造との直接の相互作用によって、松果体のはたらきを変える。環境の光は、日周期をその周期に一致させる働きと伝達する働きをもっているとみなされ得る。内在性振動発生装置を光周期にのせるための光後下はゆっくりであるが、信号伝達に及ぼす光の後下は速やかである。そして、それによって、光によってN-アセチルトランスフェラーゼが速やかに“消失”することと、持続光によって日周期が妨げられることを、おそらくは説明できるであろう。視交叉上核からの単一神経路が、松果体によるメラトニン形成に関係する両酵素を調節しているが、これらの結合についての詳細は、なお、あきらかになっていない。間接的に証明された根拠は視交叉上核から松果体への神経路には、視床下部の灰白隆起の領域、内側前脳束および中間質外側路への中継路が含まれることを示唆している。このように、松果体は、交感神経ニューロンを経て受けた信号をメラトニンという内分泌物に変換させる、神経内分泌変換体ともいうべきものであるらしい。松果体の分泌は視床下部の働きを変化さえるが、それは、内分泌が心身の血液循環あるいは、脳脊髄液に入ってからののちに作用する。松果体のセロトニンとメラトニンの日内変動は、光入力の周期に応じて起こる。松果体活動のこれらの周期性変化は、この腺が生物学的時計としてはたらいて、生理学的過程と行動学的過程とを調節する信号を出していることを示唆している。これらの変動は日周期circadian rhythmsと呼ばれ、環境からの刺激が存在すれば、ほぼ24時間周期を示すだけである。松果体実質の腫瘍は、性的機能を低下させて思春期を送らせるが、他方、松果体を破壊するとしばしば早発思春期を伴う。これらの観察は、松果体が、性腺および生殖器系に抑制的影響を及ぼすことを示す実験的研究の結果と一致する。
19. Splenium, corporis callosi(脳梁膨大)Sphenium of corpus callosum
→脳梁幹の後端は著しく膨大化し、松果体と蓋板とを上方から被っている。膨大部と中脳との間の大脳横裂からは脳軟膜が進入し、脈絡組織を形成する。
20. Gyrus cinguli (帯状回)Cingulate gyrus
→帯状回は、脳梁の前端から始まり、脳梁の上を回って、その後端まで達する。帯状回と脳梁の間には脳梁溝があり、帯状回と上前頭回との間には帯状溝がある。
21. Lubulus paracentralis(中心傍小葉)Paracentral lobule
→中心傍小葉は中心前回と中心後回との半球の内側面への延長部で形成され、中心溝による切れ込みがある。
22. Falx cerebri(大脳鎌)Falx cerebri; Cerebral falx
→大脳鎌は大脳縦裂の中に入り、下方に刃の部を向けた鎌状をなし、前方は鶏冠にはじまり、上縁は前頭稜および上矢状洞溝に粗って後走し、内後頭隆起に付く。上縁はその中に上矢状静脈洞を含む。下縁は自由縁で、大脳縦裂の中で脳梁のやや上方を走り、その中に狭い下矢状静脈洞を含む。大脳鎌の後下縁は小脳テントの上面と癒着し、その癒着縁は直静脈洞を含む。
23. Fornix(脳弓)Fornix
→白い線維が帯状になった脳弓は海馬体の主要な遠心性線維系を構成する。この中には投射線維と交連線維の両者が含まれる。これは海馬台皮質(海馬台前部、海馬台、前海馬台)と海馬の大錐体細胞の軸索からなり、海馬白質として側脳室表面に広がり、それらがまとまって海馬采を形成する。両側の海馬采は後方へ進むにつれて太くなり、海馬の後端に至って脳梁膨大の下を脳弓脚となって子を描いて上がると同時に両側の物が互いに近付いてくる。このあたりで多数の線維が反対側の脳弓に入る。すなわち交叉線維が薄く板状に広がって脳弓交連(海馬交連、または脳琴psalterium)を形成するがヒトでは発達が悪い。両側の脚は合して脳弓体となり脳梁の直下を前方に視床の吻側端まで行き、ここで再び線維束が左右に分かれ脳弓の前柱として室間孔から前交連の後ろまで腹方に曲がる。神経線維が薄い帯状になった海馬采は脳弓のほぼ全経過にわたって外側に位置しているが、吻側では脳弓の本体である脳弓前柱の中に混ざってしまう。脳弓線維の最大部分は前交連の尾側を交連後脳弓として下行し、残りは前交連の前を交連前脳弓となって走る。海馬台に起始をもつ交連後脳弓線維は視床下部を通過して乳頭体に至るが途中視床にも枝をだす。乳頭体では主として内側核に終止する。視床下部の吻側部で交連後脳弓から分かれた線維は、外側中隔核と、視床の前核群や外側背側核に終わる。その他、海馬台からの遠心線維は、前頭葉内側皮質、尾側帯状回、海馬傍回などに直接投射する。交連後脳弓線維の一部は乳頭体を越えてさらに尾方へ下り中脳被蓋に入る。交連前脳弓は線維束としては交連後脳弓よりも小さく、疎であって肉眼的に認められるようなものではない。これらの線維は海馬のすべてのセクターの錐体細胞に始まり、中隔核群の尾部の一部に終わる。以上にのべた解剖学的な連絡様式から示唆されるように、海馬台からのインパルスは複雑な経路を経て種々の部位に投射される。すなわち、海馬と海馬台は直接あるいは関節の投射路によって中隔核群、視床下部、視床、大脳皮質の広汎な領域および中脳網様体と結合する。
最終更新日: 19/02/05