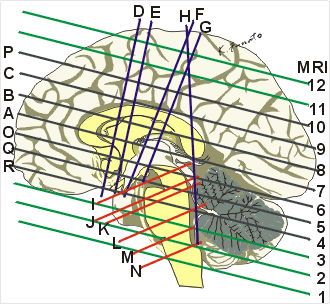
30歳♂のT1強調MRI画像
外眼角耳孔面に水平な面(線条体[被殻と尾状核]、内包、視床下部ならびに視床の腹側部が見える)
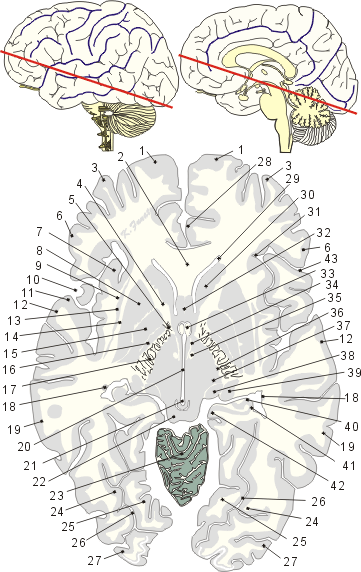
01:上前頭回、02:脳梁膝、03:中前頭回、04:内包前脚、05:内包膝、06:下前頭回、07:島、島葉、08:被殻、09:最外包、10:外側溝後枝、11:側頭葉、12:上側頭回、13:前障、14:外包、15:淡蒼球内節の外側部、16:淡蒼球内節の内側部、17:内包後脚、18:尾状核尾、19:中側頭回、20:第三脳室、21:中脳水道、22:下丘、23:小脳虫部、24:外側後頭側頭回、25:内側後頭側頭回、26:側副溝、27:後頭葉の大脳回、28:帯状回、29:側脳室前角、30:尾状核頭、31:島輪状溝、32:透明中隔、33:脳弓柱、34:視床下部、35:乳頭視床束(ブィック・ダジール束)、36:視床網様核、37:視床後外側腹側核(外腹側後核)、38:内側膝状体、39:外側膝状体、40:海馬白板、41:海馬、42:海馬傍回、43:外側溝上行枝01. Gyrus frontalis superior(上前頭回)
→上前頭回は上前頭溝の上にある。上前頭回および中前頭回の後部には運動性皮質中枢の続き(運動前野)があるが、これらの回の前部および下前頭回は連合中枢(前頭連合野)と考えられれる。
02.Genu corporis callosi(脳梁膝)Genu of corpus callosum
→脳梁膝は脳梁吻の上方に位置し、線維は前方に向かって走り、前頭葉に放散している(小鉗子)。
03.Gyrus frontalis medius(中前頭回)Middle frontal gyrus
→中前頭回は上前頭溝と下前頭溝の間にあり、中前頭回および下前頭回は人脳では特に発育が良い。
04.Crus anterius capsulae internae(内包前脚)Anterior limb of internal capsule
→内包前脚はレンズ核と尾状核頭部との間にある。内包前脚には、前視床縫線、前頭前皮質橋核路が含まれる。
05.Genu capsulae internae(内包膝)Genu of internal capusle
→内包膝は前および後脚の間にある。皮質延髄路および皮質網様体路の線維が含まれる。
06.Gyrus frontalis inferior(下前頭回)Infeiror frontal gyrus
→下前頭回は外側溝の前枝と上行枝によって3つの部分に分けられる。すなわち、①眼窩部、②三角部、③弁蓋部である。優位大脳半球(通常は、右利きの人では左側にある)の三角部と弁蓋部は運動性言語中枢motor speech center(ブローカ中枢Broca's area)があり、言語活動に必要な微妙な運動支配を支配すると言われる。前頭葉下面は前頭骨眼窩部の上面にのっておりわずかに凹んでいる。ブローカ中枢は、右利きの人では左半球に、左利きの人では右半球にあるという説がある。ブローカ中枢が侵されると、他人の話を聞いて理解できても自分から話すことができない状態(運動性失語症motor aphasiaまたは表現失語expressive aphasia)になる。
07.Insula; Lobus insularis(島、島葉)Insula; Insular lobe
→ライルの島とも呼ばれる。外側溝の深部にある大脳皮質で、その表面は前頭葉、頭頂葉および側頭葉によりおおわれている。島をおおっているこれらの大脳葉の部分を前頭弁蓋、前頭頭頂弁蓋、および側頭弁蓋という。すなわち、弁蓋は島をおおう外套部である。島の周囲は輪状溝により囲まれ、これにより弁蓋と境される。この溝は島の下端部では欠き、この部分を島限とよぶ。ほぼ胎生17週からこの部はその周囲が厚くなるため、陥没してその輪郭が明瞭になり、第19週ごろから前頭葉、側頭葉および頭頂葉の発達につれてしだいにこれらによっておおわれ、生後は全くかくれてしまう。このように島の表面には前頭葉、頭頂葉、および側頭葉の部分が延びて来て、外側溝後枝の上下唇をなし、島を被っている。島は後上方から前下方に走る島中心溝によって後方の1~2個の長回と前方の4~5個の短回に分かたれる。Reil, Johann Christian (1759-1813)オランダ人解剖学者。精神病理学者。大脳のライル島を記述("Exercitationum anatomicarum fasciculus primus.etc", 1796)、生体の生理学機能の、化学的表現としての生命力を提唱("Von der Lebenskraft", Arch. Physiol, (Halle), 1796, 1,8-162)。最初の生理学雑誌"Arch. Physiol."と最初の精神病学雑誌"Magazin fur Nerven heilkundle"を刊行。
08.Putamen(被殻)Putamen
→被殻はレンズ核の外側部を形成し、外側髄板によって淡蒼球の外節とへだてられている。島皮質とは最外包、前障、外包によってわけられる。被殻の構造は尾状核とまったく同様で、太い有髄線維をほとんど含まず、主として小さい神経細胞からなるが、散在性の大細胞を含む。被殻と尾状核は発生学的にみると、同一の細胞群が内包の発達によって隔てられたもので、両者の間には互いに結合する灰白質の線条が多数見られる。そのため、両者をあわせて線条体または新線条体と呼ぶ。線維連絡も尾状核と原則的に等しい。霊長類において動物が高等になると、相対的な意味で尾状核の体積が減少し、被殻の体積が増大するといわれている。
09.Capsula extrema(最外包)Extreme capsule
→前障の外側方には幅の狭い線維層があって、島の皮質と前障を隔てている。これらの線維の連絡関係は知られていない。
10.Ramus posterior sulci lateralis cerebri(外側溝後枝)Posterior ramus of lateral cerebral sulcus
→外側溝の後枝は縁上回へ終わる長い枝。
11.Lobus temporalis(側頭葉)Temporal lobe
→側頭葉は外側溝より下方にある部分で、上外側面から下面におよび、後方は後頭葉および頭頂葉に移行する。上外側面では溝としては前後に走る上および下側頭溝があり、これらによって上、中および下側頭回が区画される。上側頭回の背側面で外側溝にかくれた部分には3本の横側頭溝があり、これらにより区別される二つの横側頭回(Heschl)がある。ここに聴覚野がある。これらの大部分は側頭葉下面皮質とともに連合中枢(側頭連合野)と目される。優位半球(主に左脳)の上側頭回の後部から角回にかけて感覚性言語中枢(Wernicke野)があるとされる。側頭葉下面では溝として前後に走る後頭側頭溝およびその内側をほぼこれと平行に走る側副溝がある。また側副溝の前方の延長部には浅い嗅脳溝がある。これは海馬傍回の前部と側頭葉の残部とを境する。回としては外側後頭側頭回は大脳上外側面における下側頭回が下面へ直接移行したもので、これらの両回の間を境する溝はない。内側後頭側頭回は後頭側頭溝と側副溝の間にあり、舌状回は側副溝後部と鳥距溝の間にあり、むしろ大部分は後頭葉に属する。
12.Gyrus temporalis superior(上側頭回)Superior temporal gyrus
→上側頭回は外側溝と上側頭溝の間にある。上側頭回の後部付近には感覚性言語中枢sensory speech center(ウェルニッケ中枢Wernicke's area)があり、これは聞いた言葉を理解する中枢であるという。一次聴覚野は聴覚の中枢で、側頭葉の上側頭回の上面(Brodmannの41野)にある。二次聴覚野は一次聴覚野の周囲(42、22野)にある。一次聴覚野で聞く音の意味はこの皮質領域で理解される。
13.Claustrum(前障)Claustrum
→前障はレンズ核と島との間にある、内側が凹面をなす板状の核で、腹側方に厚くなる。この核とレンズ核との間には外包があり、また島の皮質との間には最外包がある。これらは狭い白質で、大部分は連合線維から、一部は交連および投射線維からなる。前障は種々の視床核、扁桃体などから線維を受け、大脳皮質に広く投射する。前障は以前は線条体とともにいわゆる基底核に数えられたり、あるいは皮質層の付け足しとして島皮質に属するものとされた。しかしながら、発生学的ならびに比較解剖学的研究によって、前障は発生の途中で位置がずれた古皮質の細胞群であることが証明されている。前障はその広い底の所で古皮質の領域へ移行する(すなわち梨状前野や扁桃体の外側核へ)。頭頂葉、側頭葉および後頭葉の皮質からの、無髄線維が局在的配列をなして前障に終わると言われている。前障の機能についてはわかっていない。
14.Capsula externa(外包)External capsule
→前障と被殻の外表面とのあいだには幅の狭い白質の層があり、大脳皮質から被殻に達する神経線維はこの線維層を通る。
15.Globus pallidus lateralis(淡蒼球内節の外側部)Lateral part of globus pallidus
→淡蒼球の外側髄板と内側髄板にはさまれた部分。
16.Pars medialis globi pallidi(淡蒼球内節の内側部)Medial part of globus pallidus
→淡蒼球の内側髄板内側方にある部分。
17.Crus posterius capsulae internae(内包後脚)Posterior limb of internal capsule
→内包後脚の主部は大部分視床とレンズ核の間の空間を占め、ほぼ垂直の方向に走る線維群からなる。その前部に皮質脊髄線維(頭部を除く上半身に対する線維は前、下半身に対するものは後)があり、皮質脊髄線維の外側を皮質赤核路が走る。皮質脊髄線維の後方には上視床脚(視床後腹側核から大脳皮質の知覚野に至る線維および前外側腹側核から運動野と運動前野に行く線維を含む)がある。その後方に、レンズ核の後方をほぼ水平に後頭葉に向かう部分がある。これは後脚のレンズ後部で後視床脚、視放線、頭頂橋路、皮質被蓋路などがここを通る。後脚のレンズ下部はレンズ核の腹側を外側方に延びて側頭葉に向かう部分で、下視床脚、視放線および側頭橋路を含む。系統発生学的に古い投射路は古皮質および原始皮質から起こり、通常内包を通らず皮質かの脳部に至るもので、そのうち特異な所見を示すのは脳弓である
18.Cauda nuclei caudati(尾状核尾)Tail of caudate nucleus
→尾状核尾は視床と分界条によって境される。側脳室の中心部の底面の外側縁を後走し、ついで側脳室の弯曲に従って前下方に開いた弓を描きつつ下角の上壁に至り、扁桃体の後端部の外側部に接して終わる。
19.Gyrus temporalis medius(中側頭回)Middle temporal gyrus
→側頭葉には大脳回が3列に並んでいる。つまり中央の上側頭溝と下側頭溝に挟まれた脳回が中側頭回である。
20.Ventriculus tertius(第三脳室)Third ventricle
→左右の間脳の間にある背腹方向にスリット状を示す腔である。前壁は終板と前交連によってつくられる。前上部には室間孔が開口し、左右の側脳室と交通し、後方は中脳水道と連絡する。後壁は松果体に入り込む松果体陥凹がみられ、下壁は視床下部によってつくられ、視交叉陥凹、漏斗陥凹がみられる。外側壁を形成している視床と視床下部の境には視床下溝が走る。なお、脳室の前上方部に第三脳室終脳部とよばれる部分がある。
21.Aqueductus mesencephali; Aqueductus cerebri(中脳水道)Aqueduct of midbrain; Cerebral aqueduct
→『シルビウス水道』ともよばれる。中脳では脳室系は細い管となり、間脳の第三脳室と菱脳の第四脳室とを結合する。これを中脳水道と称し、横断面は円形または底辺を背側に向けた角のとれた三角形をなし、中心灰白質によってかこまれる。その存在については古くから知られていたが、フランスの解剖学者Jacobus Sylvius (1478-1555)の著書(1555年)で初めて説明がなされた。
22.Colliculus inferior(下丘)Inferior colliculus
→下丘は中脳蓋を形成する二対の隆起(四丘体)のうち下方の一対をいう。下丘は聴覚系の中脳における中継核で、細胞構築および機能的に中心核、外側核および周囲核の三つの核からなる。下丘核は外側毛帯を介して蝸牛神経核および台形体核から線維を受け、下丘腕を通って両側性に視床の内側膝状体へ線維を送る。
23.Vermis cerebelli (小脳虫部)Vermis of cerebellum
→小脳虫部は小脳のなかで系統発生学的に古い無対の部分。
24.Gyrus occipitotemporalis latealis(外側後頭側頭回)Lateral occipitotemporal gyrus
→後頭側頭溝の外側にある回。側頭葉下縁で切れ目なく下側頭回へ移行する。
25.Gyrus occipitotemporalis lateralis(内側後頭側頭回)Medial occipitotemporal gyrus
→内側後頭側頭回は後頭極から側頭極まで延びている。この脳回の内側縁は側副溝と嗅溝によって境され、外側縁は後頭側頭溝によって境されている。
26.Sulcus collateralis(側副溝)Collateral sulcus
→側頭葉の側頭極からはじまり、海馬傍回と内側後頭側頭回の間にある溝。後頭葉まで達する。
27.Gyri occipitales(後頭葉の大脳回)Occipital gyri
→後頭葉の大脳回を一括して後頭葉の大脳回と呼んでいる。
28.Gyrus cinguli(帯状回)Cingulate gyrus
→帯状回は、脳梁の前端から始まり、脳梁の上を回って、その後端まで達する。帯状回と脳梁の間には脳梁溝があり、帯状回と上前頭回との間には帯状溝がある。
29.Cornu frontale; Cornu anterius(側脳室の前頭角、側脳室の前角)Frontal horn; Anterior horn
→室間孔より前方。内側は透明中隔、外側は尾状核頭、上方は脳梁幹、前方は脳梁膝または脳梁吻で境される。
30.Caput [nuclei caudati](尾状核頭)Head of caudate nucelus
→尾状核頭は側脳室の前角の中に膨隆し、その外側壁をなし、視床の前方に位置する。尾状核頭は後方にしだいに小さくなり、視床の出現とともにその背外側に位置するようになり、尾状核尾に移行する。
31.Sulcus circularis insulae(島輪状溝)Circular sulcus of insula
→島の境界となる溝。島限で中断する。
32.Septum pellucidum(透明中隔)Septum pellucidum
→透明中隔は左右の側脳室前角を分離する一対の薄板である透明中隔板と、その間の狭い間隙である透明中隔腔からなる。透明中隔腔は成人ではしばしば閉鎖し、左右の透明中隔板が密着する。透明中隔腔は脳室ではなく、その内面には上皮細胞層が証明されない。透明中隔板は脳梁と脳弓の間に張られているが、発生学的には終脳胞の内側面の一部が脳梁の発達のため前頭葉から分離されたもので、痕跡的な大脳皮質の構造を示す。透明中隔は元来の中隔野の後部の一部で、中隔野には透明中隔のほか、前交連と終板の前にある終板傍回、梁下野、中隔核などが含まれる。
33.Columna fornicis(脳弓柱)Column of fornix
→脳弓柱は一部は第三脳室壁中にある脳弓の前部。乳頭体まで達す。
34.hypothalamus(視床下部)Hypothalamus
→視床下部は間脳の中で、内臓機能、自律機能および内分泌機能と最も関係が深い部分である。これらの機能のすべてが、感情的および情動的行動と密接に関係している。第三脳室の側壁の下部および底にあたる。脳底面からみると、吻側から数えて、視交叉、漏斗、灰白隆起、乳頭体とつづき、漏斗の先端は下垂体に連なる。背側は視床下溝により視床と境されており、吻側は終脳の視索前野に、尾側は中脳被蓋と中脳中心灰白質に、尾外側は視床腹側部に移行する。通常、矢状面に平行な三つの帯状領域、すなわち、視床下部脳室周囲層、視床下部内側野、視床下部外側野に区分される。これらの間を多数の細かい神経線維が主として吻尾方向に走っている。視床下部は前後径が約10mmである。視床下部は内部環境を正常に維持する機序に関与し、また心悸亢進、瞳孔散大、“冷汗”の分泌などの情動反応の表出にも一役を演じている。大脳皮質を除去し、背側視床を除去した後でも、怒り反応はあらわれる(「みかけの怒りSham rage」)。さらに、視床下部は成長、性的成熟など新地あの成熟過程にも関与している。したがって、視床下部を損傷すると、広汎で顕著な内分泌性、代謝性、行動性(情動性)の以上が一緒に起こってくることになる。視床下部は下垂体ホルモンによって内部環境に影響を及ぼすが、さらに脳幹網様体や自律神経系を介しても同様の働きを示す。視床下部への情報は通常の入力神経系によって伝達されるばかりでなく、視床下部のニューロンは内部環境からの物理的および化学的刺激(血液の温度など)にも反応する。植物性機能の中には、その統御中枢が視床下部自体に存在するものもあるが、呼吸や心臓の活動などの機能に関しては、視床下部はもっと回の中枢に対する修飾器として働いている。
35.Fasciculus mamillothalamicus(乳頭視床束、ブィック・ダジール束)Mamillothalamic tract
→乳頭体と視床前核を結ぶ神経線維束。主乳頭束として、乳頭被蓋束とともに乳頭体の背側よりでる。乳頭体内側核からおこる神経線維は同側の視床前核のうちでも前腹束核と前内側核に分布し、乳頭体外側核からおこる神経線維は両側の視床前背側核に分布するといわれる。また、乳頭視床束には視床前核から乳頭立ちに向かう神経線維も少数含まれるらしい。
36.Nucleus reticularis thalami(視床網様核)Reticular nucleus of thelamus
→視床網様核は、外髄板と内方の間にある薄い神経核である。多くの神経線維の中に神経細胞が分布しているので網様核という名前が付けられているが、網様体とは関係がない。この核への求心性線維は、視床皮質線維と皮質視床線維の側枝である。網様核の神経細胞は、視床の神経核と相互連絡を持っており、この神経核の細胞の軸索は、自分に側枝をだしている視床の神経核に終止している。大脳皮質へは投射していない。
37.Nucleus ventralis posterolateralis(視床後外側腹側核)Ventral posterolateral nucleus of thalamus
→内側毛帯と脊髄視床路の線維がシナプス結合する。内側毛帯線維の軸索終末の分枝は境界の明瞭な小円錐状を成して視床ニューロンとシナプス結合するが、脊髄視床路線維の軸索終末は散在性に分枝知る。入力情報はニューロンを乗り換えて大脳皮質の3、1、2野の下部へ投射する。
38.Corpus geniculatum mediale(内側膝状体)Medial geniculate body
→下丘と連絡する。聴覚路の一部をなす。内側膝状体は腹側核、背側核、巨大細胞内側核に区分される。
38a. Nucleus ventralis corporis genigulati medialis(内側膝状体腹側核、内側膝状体主腹側核)Ventral principal nucleus of medial geniculate body
→内側膝状体腹側核は聴覚系「核部」の中継核であり、下丘中心核から入力線維を受け、側頭弁蓋にある一次聴覚皮質投射線維を送る。内側膝状体腹側核と一次聴覚皮質には音の周波数対応配列(tonotopic representation)がみられ、形態的には入力線維と中継ニューロンが層的構成をとって配列する。
38b. Nucleus dorsalis corporis geniculati lateralis(内側膝状体背側核)Dorsal nucleus of medial geniculate body
→内側膝状体背側核は下丘中心周囲核(pericentral nucleus)から起こる投射線維をうける。内側膝状体背側核は側頭平面と上側頭回にある聴覚連合皮質と連絡する。内側膝状体大細胞部は内層毛帯、脊髄視床路などからの側枝を受ける。内側膝状体核大細胞部は聴覚系「殻部」の中継核であるとともに体性感覚性入力などをも受けており、その他の視床非特殊核と同様、大脳皮質Ⅰ層に投射線維を送る。
38c. Nucleus medialis magnocellularis corporis gemicalati medialis(内側膝状体巨大細胞内側核、内側膝状体核の大細胞部)Medial magnocellular nucleus of medial geniculate body
→内側膝状体核の大細胞部は内側膝状体の内側部を占め、腹側核と内側毛帯の間に位置する。
39.Corpus geniculatum laterale(外側膝状体)Lateral geniculate body
→外側膝状体は、視覚系における視床の中継核であり、内側膝状体の吻側外側方、大脳脚の外側方で、視床枕の副側方にある。この核には、細胞が層状に配列した構造があり、横断切片では、門を腹内側方に向けた馬蹄形をしている。視索の視交叉および非交叉性線維は、この門を通ってはいり、一定の正確な様式に従って分布する。ヒトと霊長類では、外側膝状体を構成するものは、6つの細胞層であり、これらは2つに大別される。同心円状に並んだ6つの細胞層は、介在する線維帯によって区切られ、寒冷では、腹内側方の門の領域から始めて、1から6まで番号が付けられている。外側膝状体の背側核を区分すると、大細胞性部(1と2層)および小細胞性部(3~6層)となる。外側膝状核の背側部の2つの領域は網膜の神経節細胞から求心線維を受ける。外側膝状体の小細胞性の層を構成するのは、腹背の方向に順に3,4,5および6層であり、容易に区別される。これらの諸層を外側方にたどると、4層が6層と、また、3層が5層というように対をなす2層が1組になって癒合する。網膜から外側膝状体への投射は正確で、視索の交叉性線維と非交叉性線維とはそれぞれ、別々の層に終わる。すなわち、交叉性線維は1,4および6層に終わり、一方、非交叉性線維は2,3および5層に終わる。交叉性網膜膝状体線維関係のある2つの特殊な性状が構造に反映されている。単眼性の視野は半月形であるが、これは反対側の網膜の内側半の最も内部にある受容要素によって受け取られる。網膜のこの部分にある神経節細胞は、反対側の外側膝状体の二重層の部分に交叉性に線維を送る。この二重層は、4層と6層の部分が外側方で癒合して出来ている。網膜の内側半の中にあって、視神経線維が通っている視神経円板(乳頭)には光受容器がなく、[周辺]視野測定[法]で見つけることができる盲点の原因となる。視神経円板(乳頭)は反対側の外側膝状体の中で、4層と6層の中の細胞層が不連続である部位に対応する。ヒトの外側膝状体を通る切断切片のニッスル標本によって、細胞の直線的配列が明らかにされており、細胞の長軸の方向は、各細胞層の軸に垂直である。小細胞性の諸層にある各周囲部は、“投射の方向線”に平行であり、これらの視野中の各店点が同じように復元されていることを示している。外側膝状体の中における網膜表面との局所的対応関係は、高度に組織化され、しかも正確である。両眼視による視野の反対側半は、交叉性線維と非交叉性線維とが異なる層に終わってはいるが、外側膝状体のすべての層に投影される。6つの層における投射の場所は、完全にきまっているので、両眼視における反対側の眼の視野の中のどんなに小さな領域でも、6つの層全体を通して、“投射の方向線”に平行して放射状に延びる背腹方向の細胞柱に一致するkとを示すことができる。外側膝状体を構成するものは、馬蹄形に曲がった6つの薄い細胞層である。しかし、そこに投射してくる場所は正確にきまっている。それゆえ、“投射の方向線”の中の細胞柱は、両眼視の視野のうち反対側の視野に関係する各眼の網膜の中の対応する点からの入力を受ける。両眼視の像の融合は、外側膝状体の中では起こらない。それは、網膜膝状体線維が外側膝状体の異なった層に終わっているからである。(術後長期間生存させた例の)視神経の切片を追って調べると、順応性の変性、またはニューロン越えの変性は、各側の外側膝状体の3つの層に起こる。神経線維、または細胞の変性が起こる層は、網膜からの交叉性線維(1,4および6層)と非交叉性線維(2,3および5層)の配列に従って異なる。網膜の小さい傷によって、同側、対側の3つの違った層の中に“投射の方向線”に従って、一列に並んで配列した細胞集団に、ニューロン越えの変性が起こる。反対側の単眼視野(単眼性の半月形の視野)は、網膜の内側半のもっとも内側の部分における受容器要素と関係しており、これからは、2層性の部分に終止する交叉性の神経線維のみが出ている。外側膝状体における“投射の方向線”は、有線野に加えた傷害によって外側膝状体の中に現れる逆行性細胞変性の研究からも、明らかにすることができる。視野の半分は、局在性をもってそれぞれの側の半球の有線野に部位局在的に投射しているので、外側膝状体の中に逆行性細胞変性が現れる帯状の部分は、それぞれの側で、投射の方向線によって境される。外側膝状体では視野の中心を通る水平線が背腹方向に入る斜めの面に対応し、この面により内側部と外側部が分けられる。両眼の網膜の上半部からの神経線維は、外側膝状体の内側半部に投射し、下半分は、本核の外側半部に線維を送る。網膜の黄斑からの投射は、外側膝状体尾側部のうち視野の中心を水平子午線に相当する面の両側にある。幅の広いクサビ形の部分として示される。黄斑に相当する部分は、外側膝状体の全容積のおよそ12%ヲ占める。視野の中心を通る垂直の子午線に一致し、本核の尾側縁に沿って、内側の境界からの外側境界に及んでいる。外側膝状体は、視索の主要な終止場所である。ここから膝状体鳥距路、あるいは視放線を経て、鳥距周囲皮質(17野)に投射があり、また外側膝状体は、この皮質野から皮質膝状体線維を受ける。本核は、視床枕と核間結合をしている。
40.Alveus hippocampi(海馬白板)Alveus
→海馬白板は有髄線維の薄い層であり、側脳室の上衣細胞層に直接面している。白板の線維のなかには海馬からの出力線維(錐体細胞の軸索)も、海馬への入力線維も存在する。
41.Hippocampus(海馬)Hippocampus
→海馬は(海馬はウマの前半身と長い魚の尾をもったギリシャ神話における動物)またはアンモン角(アンモンはエジプトの巻いた角をもった神)は海馬溝による長い弯曲したたかまりで、側脳室下角の内側壁の大部分を占め、その肥厚した前端部の上面にはいくつかの指のような肥厚があり、これは海馬足または海馬指とよばれ、ほとんど下角の前端に達している。海馬の側脳室表面は海馬白板によっておおわれる。これは内側方は海馬采に続く。海馬采は海馬をおおってその内側部に付く。大脳半球内側面の部分から形成される古い皮質(原皮質)で解剖学的に海馬を厳密に定義することはむずかしい。すでに爬虫類で海馬に相当する皮質がみられ、哺乳類でみられる基本的な要素が発達している。ある種の出力と入力が共通しており、互いに密接に関連(結合)している単純な層形成をもっている部分、すなわち、固有の海馬(アンモン角)と歯状回および、ときに海馬台(海馬支脚、海馬床)、海馬采、脳弓を加えて考えた方が研究目的上都合がよいことが多い。また、しばしばこの意味で、海馬形成(hippocampal formation)という言葉が用いられる。さらに皮質分野27(海馬支脚前野),29e(脳梁膨大後野)、49(海馬支脚傍野),28(内嗅領),からなるretrohippocampal foramtionを含めて海馬領域(Angevin, 1965)とよばれる。海馬は構造上部位差がみられ、CA1~CA4の亜核に分けられる(Lorente de No, 1933, 1934)。また、構造上、分子層、錐体細胞層および多形細胞層の各層に区別される。海馬からの遠心性線維は、主として前部から中核部に、また、主として後部から脳弓線維となって視床下部とくに乳頭体に終わる。求心性線維には、嗅内野(entorhinal area, 28野)や嗅周野からくる穿通線維および中隔核や対角帯核からくる中隔海馬線維がある。嗅脳からの直接投射はない。海馬は以前、嗅覚系に関係すると考えられ、ついで情動に、最近では臨床的知見により記憶、とくに短期記憶に関係があることが示唆している。側頭葉基底部の皮質および海馬を両側性に除去すると最近のできごとに関係する記憶が失われる。患者はまったく正常に会話についていけるが、話題が変わるとたちまち前の話の筋を忘れてしまう。
42.Gyrus parahippocampalis(海馬傍回)Parahippocampla gyrus
→海馬傍回は海馬溝の下方、側副溝との間に大きな回である。海馬傍回の皮質の細胞構築は海馬台前部から順次海馬台、前海馬台を経て海馬体、歯状回にいたる間に次第に6層から3層構造に移り変わる。嗅内領(28野)は6層構造の皮質であるが、より内側ではある層が脱落して再構成が行われる。海馬体皮質には基本的な多形細胞層、錐体細胞層、分子層よりなる3層構造がある。これらの細胞の軸索、樹状突起の配列によりいくつかの層が二次的にできる。錐体細胞の軸索は海馬白板に投射し脳弓の海馬采となる。
43.Ramus ascendens(外側溝上行枝)Ascending ramus of lateral cerebral sulcus
→外側溝の上行枝は前頭葉中にある外側溝の上行する短い枝。
最終更新日: 19/02/05