| 脳神経の概要 |
| 終神経[0] |
| 嗅神経[I] |
| 視神経[II] |
| 動眼神経[III] |
| 滑車神経[IV] |
| 三叉神経[V] |
| 外転神経[VI] |
| 顔面神経[VII] |
| 内耳神経[VIII] |
| 舌咽神経[IX] |
| 迷走神経[X] |
| 副神経[XI] |
| 舌下神経[XII] |
|
最終更新日: 12/04/24
|

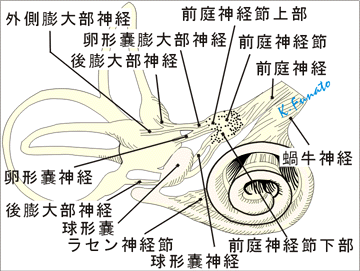
内耳神経[Ⅷ]
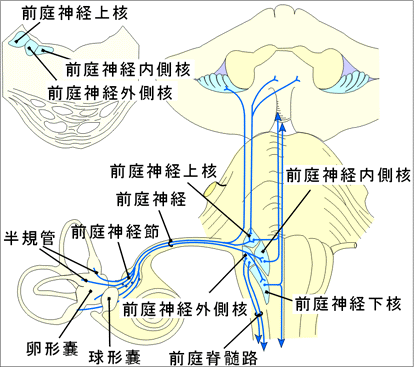
内耳神経は蝸牛神経と前庭神経とからなり、それぞれ蝸牛管(聴覚)と前庭(平衡覚)の感覚上皮から来る刺激を、菱形窩にある蝸牛神経核と前庭神経核につたえる。蝸牛神経と前庭神経は内耳の感覚上皮から起こり、前者は蝸牛軸にあるラセン神経節から内耳道底のラセン孔列に達し、後者は内耳道底の前庭神経説に達する。内耳道底で蝸牛神経と前庭神経は合し、内耳神経に達する。内耳道底で蝸牛神経と前庭神経は合し、内耳神経となって内耳道を通り、橋の後縁で、顔面神経の外側から脳へ入る。 前庭神経は系統発生的には蝸牛神経よりも古く。固有受容感覚と相補的な関係にある。蝸牛神経は外受容感覚に関与する。両神経の作用はまったく異なるようにみえるが、平衡感覚(前庭、半規管)や蝸牛における管状構造および液体の存在、その感覚上皮が両者とも液体新藤を受容する有毛細胞であることを考えると共通の点が多い。なお、蝸牛神経では大脳皮質に至る求心性投射路の発育が良好であるが、前庭神経の求心性投射路は主として小脳(虫部)で終わり、小脳経路以外の大脳皮質に至る投射路は明らかでない。
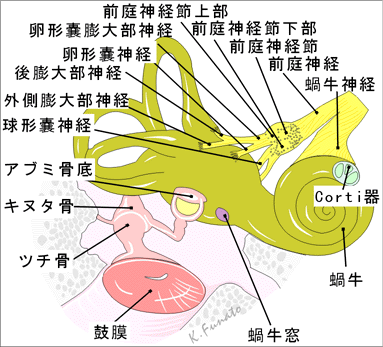
1.内耳
A.迷路内耳神経の末梢受容器は液体を容れて複雑に連なっている上皮性の管の中に存在し、この管は側頭骨岩様部内部の骨迷路におさまっている。前庭神経の受容器は半規管、卵形嚢、球形嚢の特殊化した部分に存在する。蝸牛神経の受容器は蝸牛の中のCorti器官にある。 三つの半規管はそれぞれが直角の面に配列されている。外側(水平)半規管は頭を前方に30度傾けると水平面に位置するようになる。右の前(垂直)半規管のそれと平行であり、また、左の前半器官が存在する面と右の後半器官が存在する面とは平行である。 B.骨迷路内耳の骨迷路は、蝸牛、前庭、半規管の3部からなる。骨迷路は側頭骨岩様部の外側部のかなり部分をしめる。
①蝸牛骨迷路の貝殻形の部分は、蝸牛管という内耳の聴覚に冠する部分を含む。蝸牛は、蝸牛軸という骨の芯の周りを2回転半し、蝸牛軸には血管と神経のための管がある。鼓室の内側壁に岬角をつくるのは、蝸牛基底部の大きな回転である。
②前庭小さな楕円形の小室(長さ約5mm)は、卵形嚢と球形嚢という平衡装置の部分を含む。前庭は前方で骨性蝸牛とつながり、後方で半規管とつながり、後頭蓋窩と前庭水管でつながらる。この水管は側頭骨岩様部の後表面に伸び、内耳道の後外側に開く。ここには内リンパ管と2本の小血管が含まれる。
③骨半規管この管(前・後・外側)は骨迷路の前庭と交通する。管は前庭の後上方にあってここに開き、3本が互いに垂直な位置にある。管は管は三次元空間の3つの平面をなす。各半規管は円のほぼ2/3をなし、直径約1.5mmであるが、一端には膨大部と呼ばれる膨らみがある。骨半規管の前庭への開口は5つしかないが、それは前と後ろの骨半規管が1本の脚を共有するからである。骨半規管の中には膜半規管がある。 C.膜迷路膜迷路は互いにつながる嚢と膜からなり、骨迷路の中に浮かんでいる。膜名は内リンパという水様液を含むが、その成分は膜迷路の外側で骨迷路の中にある外リンパとは異なる。膜迷路は大きく3部からなる。
・卵形嚢と球形嚢は互いにつながる小さな2つの袋で、骨迷路の前庭の中にある。 ・3つの膜半規管は骨半規管の中にある。 ・蝸牛管は蝸牛の中にある。
膜迷路骨迷路の中に浮かんでいる。蝸牛の内面を覆う骨性のラセン状の肥厚であるラセン靱帯は、蝸牛管を蝸牛の中に安定させる。膜迷路のさまざまな部分の嚢と管が、互いにつながって閉じた系をなす。膜半規管は5つの開口を通して卵形嚢に開き、卵形嚢と球形嚢は連嚢管によってつながり、ここから内リンパ管が起こる。球形嚢と蝸牛管は、結合間としてしられる細い交通を介してつながる。 卵形嚢と球形嚢にはどちらも、斑と呼ばれる感覚上皮の特殊領域がある。卵形嚢斑は卵形嚢の床にあり、頭蓋底と平行だが、球形嚢斑は球形嚢の内側壁にあり、垂直に位置する。斑の有毛細胞は、内耳神経(Ⅷ)の枝である前庭神経によって支配される。一次感覚ニューロンは前庭神経説にあり、内耳道の中にある。内リンパ管は後頭蓋窩の骨を貫き、内リンパ嚢と呼ばれる拡張部となって終わる。これは側頭骨の岩様部の後面をおおう硬膜の下にある。内リンパ嚢は、膜迷路の毛細血管からつくられる過剰な内リンパを蓄える容器である。 膜半規管のそれぞれには、一端で膨大部となって拡がる膨大部稜という感覚領域がある。膨大部稜は、管の平面内での頭の回転によって起こる膨大部内の動きを記憶する感覚装置である。膨大部稜の有毛細胞は、斑の有毛細胞と同様に、前庭神経節に細胞体のある一次感覚ニューロンにより支配される。 蝸牛管はラセン状の盲管で、ラセン靱帯によって蝸牛の内側壁と外側壁のあいだにある。蝸牛管の天井は前庭膜がつくり、床は基底膜と骨ラセン板の外側部がつくる。聴覚刺激の受容器はラセン器(Corti器)で、基底膜の上に乗っている。その上にはゼラチン状の蓋膜が被さっている。ラセン器には有毛細胞があり、音波によって生じた内リンパの振動に反応する。
D.内耳道内耳道は、層橈骨の錐体の中を外側にむかって約1cmほど走る細い管である。内耳道の開口は錐体の後内側面にあり、外耳道の延長にある。外耳道の外側部は孔のあいた薄い骨板で閉じられ、内耳と隔てられている。この骨板を、顔面神経(Ⅶ)、内耳神経(Ⅷ)の枝、血管が通り抜ける。内耳神経は内耳道の外側端近くで分岐し、蝸牛神経と前庭神経になる。
2.前庭神経
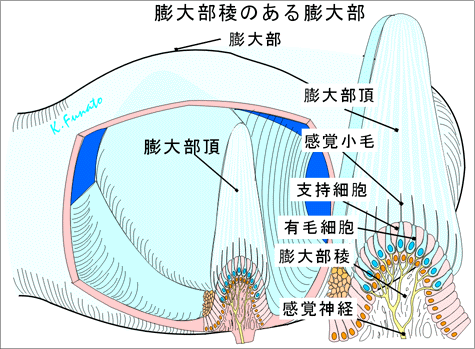
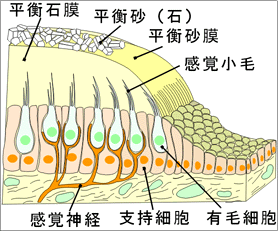
①前庭迷路の感覚上皮半規管の特殊感覚要素は管が拡大して膨大部となったところになる(膨大部稜)。三つの半規管の末端にある膨大部は同型で同サイズである。膨大部稜は管腔内に突出した一種の稜であり、稜にはゼラチン様の膨大部頂がかぶさっており、その一方の端は膨大部の反対側の管壁に達している。このように、半規管の腔は弁で閉じられたようになっている。三つの膨大部稜はそれぞれ前庭神経の枝である前・後・外側膨大部神経の支配を受けている。 感覚上皮は卵形嚢と球形嚢にもある。それらは管壁の比較的小さい部分に限局しており、卵形嚢斑および球形嚢斑と呼ばれる。これらも膨大部稜と同様ゼラチン様物質でおおわれており、その物質の中には炭酸カルシウムの結晶、すなわち平衡石の比重はまわりの内リンパよりもはるかに大きい。
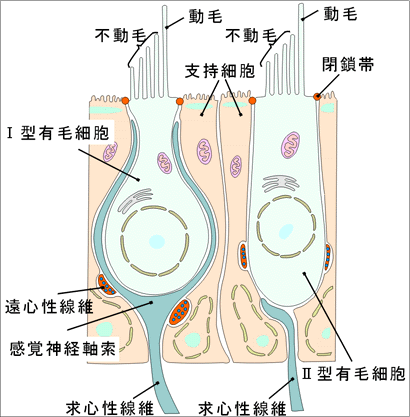
②前庭迷路の感覚上皮の構造感覚上皮にはⅠ型(ビン型)とⅡ型(円柱型)の2種類の受容細胞がある。Ⅰ型は大型の杯状の神経終末(杯)に取り囲まれており、Ⅱ型は細胞体の底部に接する小形(芽状)の神経終末に支配される。受容細胞の表面には60~100本のいわゆる不動(線)毛があり、それらが膨大部頂に埋まっている。不動毛のある細胞表面の一端には振動する運動(線)毛が1本ある。5個の特殊感覚上皮(三つの膨大部稜と二つの平衡斑)は前庭神経の6本の枝の支配を受ける。
「卵形嚢膨大部神経、卵形嚢神経、前膨大部神経、外側膨大部神経、後膨大部神経、球形嚢神経」 これらの線維は前庭神経節の双極細胞の末梢性突起であり、中枢性突起の方は前庭神経を形成する(特殊体性感覚神経)。 静止時には感覚線毛は受容細胞の線毛面に対して垂直位をとり、受容細胞は一定の静止時頻度で発火している。頭が廻ると、内リンパの慣性のために膨大部頂が曲がる。膨大部頂が曲がると線毛は切線方向に力が加わったときのように曲がる。膨大部頂がその反対方向に動くと発火頻度が減少することが知られている。 膨大部稜の線毛はすべて同じ方向の極性をもっており、回転運動にさいして、同一平面にある一対の半規管のうちの一方は興奮し、他方は抑制される。回転運動の面に垂直な方向にある半規管には運動の影響は及ばない。
③前庭迷路の機能膨大部稜は角加速時計として働く。とくに頭部の運動の方向の変化が刺激となる。三つの半規管の空間的な配列によって、三つの標準方向の角加速度を捉えることができる。各半規管の膨大部稜はそれらが位置している平面内の頭の回転にとくに反応する。反転運動の開始後約20秒で内リンパと半規管の壁は同じ速度で回転するようになり、膨大部頂はその静止位にもどる。定常運動、すなわち一定の速度と方向を保つ運動は半規管によっては受容されない。 膨大部稜には力動的な機能がある。すなわち、回転運動はおのおの三つの互いに垂直な方向の要素に分解され、各半規管がそれぞれの要素の受容に参加するのである。さらに、左右の半規管が相補的に働く。 卵形嚢と球形嚢の平衡斑は、平衡石の比重を利用して、重力の静的な効果と直線方向の加速度を捉える。たとえば、直線方向に前進加速度が加わると、平衡石は平衡斑に対して相対的にうしろに取り残され、そのために線毛が曲げられてそれが適刺激となる。卵形嚢は前後方向の加速度に感じ、球形嚢は垂直方向の加速度を捉える。
A.前庭神経核群前庭神経線維は脳幹にはいると分岐して、半規管からの線維は上行枝、卵形嚢からの線維は下行枝となる。前庭神経線維の中には下小脳脚の最内側部(傍索状体)を通って直接に小脳「片葉、小節、虫部垂の最尾側部に達するものがあるが、大部分は前庭神経核でシナプス結合する。第四脳室直下には四つの前庭神経核があり、第四脳室底からみたこの領域を前庭神経野という。前庭神経野の外側に下小脳脚がある。四つの前庭神経核は次のようである。 「前庭神経核群には以下に記す下核、内側核、上核、外側核の他に、前庭神経の根線維のあいだにある介在核やx核、y核、z核などの小さな細胞群が区別されている。ただし、z核には前庭神経線維は終糸しない。」
①前庭神経下核「下行路核・脊髄路核・Roller核」下小脳脚の線維と第四脳室底のあいだにあるかなり長い核で、尾側では次第に細くなる。核の腹外側部には大形細胞があり、ここには前庭神経は終止ししない。
②前庭神経内側核「三角核、背側核、Schwalbe核」第四脳室底で前庭神経下核の内側にある。吻側部では次第に前庭神経外側核と接着する。
③前庭神経上核「隅核、Bechterew核」第四脳室底と第四脳室外側壁のあいだ「すなわち、第四脳室の腹外側隅」にあり、上小脳脚に近く、腹方には三叉神経脊髄路核が隣接する。
④前庭神経外側核(Deiters核)前庭神経線維が脳幹に達するレベルで第四脳室底に埋もれている。大形多極性で大形顆粒状のNissl小体をもつのがこの核の細胞の特徴である。前庭神経はこの核の中央部に少数終わるだけである。
⑤前庭神経からの求心性線維前庭神経核内の求心性線維終末の分布は不規則である。四つの前庭神経核のそれぞれに前庭神経からの求心性線維を欠く部位がある。シナプス結合は大部分が小形あるいは中形の細胞と行われる。半規管からの線維は内側核の吻側部と上核に終止する。卵形嚢と球形嚢からの線維は傍索状体を通って下核と内側核の尾側に終止する。直接性前庭小脳線維は索状体(下小脳脚)を通って小脳に達し、帯状線維として胴側の片葉、小節、虫部垂「尾側部の皮質」に終止する。
⑥非前庭神経性求心性線維非前庭神経性求心性線維の主なものはCajalの間質核(内側縦束を経て)、小脳、脳幹毛様体からくる。外側核には小脳虫部(主として前葉の外側部)のPurkinje細胞の軸索が多数終わる。したがって、外側核はしばしば小脳核の細胞が腹側方向に遊走したものと見なされる。 「小脳虫部やいわゆる前庭小脳のPurkinje細胞の中には前庭神経核に軸索を送るものがあるが、Purakinje細胞の軸索の大部分は小脳核に終止するのが原則とされるから。」 片葉小節葉と室頂核からの小脳性線維は傍索状体を通って下行し、狭義の前庭神経核、すなわち、内側核、上核、下核に終止する。橋毛様体や延髄毛様体からの線維も前庭神経核とシナプする結合する。
B.前庭神経核からの遠心性線維①前庭脊髄線維前庭脊髄線維は外側および内側前庭脊髄路の2系に分かれる。外側前庭脊髄路は同側性の身体部位対応的配列をもつ線維系であって、脊髄前索の外側部を脊髄の最下部まで下行する。これらの線維群を通るインパルスにより伸筋反射が促通される。内側前庭脊髄路は前庭神経内側核および下核からの線維より成り、内側縦束を通って脊髄へ下行するが、胸髄上部以下のレベルには達しない。前庭神経核による頚筋の制御はこれらの線維によって行われる。
②前庭小脳線維前庭小脳線維は主として前庭神経下核と内側核から起こり、傍索状体を通って小脳へはいる。小脳では主として小節、片葉、虫部垂尾側部の皮質と、室頂核でシナプス結合する。
③前庭中脳線維前庭中脳線維は前庭核と眼筋を支配する神経核とを連絡する。これらの線維は主として前庭神経上核と内側核から起こる。上核からの線維は主として滑車神経核と動眼神経核に同側性に投射する。内側核からの線維は主として反対側性に滑車神経核と動眼神経核に投射する。 「外転神経核は主として同側の内側核からの線維を受ける。」 また、外側核の腹側部からは同側の動眼神経核にはいる線維がでるようである。 「外側核の腹側部からは同側の動眼神経核にはいる線維が出ることも報告されている。」
④前庭視床線維この線維系についてはまだよくわかっていない。前庭から大脳皮質への投射経路の中継核としては、生理学的実験の結果から、視床後腹側核のうち下後腹側核が考えられている。 「前庭視床線維の終止部位としては、VPIのほかに、VPM・VPLの吻側部、VPMに近接するVLの部位、などが報告されている。」
⑤前庭神経の中の遠心性線維前庭神経は少数の遠心性線維を含む。これらの線維は主として同側の延髄網様体外側部から出るようであり、前庭器の感覚細胞を抑制して末梢感覚器の感受性を減弱させる。 C.内側縦束内側縦束は上行性および下行性線維を含み、それらの線維の起始もさまざまである。延髄では内側縦束は舌下神経核の腹側で縫線の両側にあり、橋と中脳では側脳腔に近く正中線のすぐ近傍に位置している。内側縦束中の上行性線維の多くは前庭神経核、とくに上核と内側核からの線維であり、これらは眼筋を支配する神経核やCajalの間質核に終止する。 下行性線維の中には前庭神経核由来のものも、そうでないものもある。前庭神経核からの下行性線維は内側核から起こり内側前庭脊髄路を形成する。前庭神経核以外の由来をもつものにCajalの間質核からくるものがあるが、これらは間質核前庭路を形成する。視蓋や脳幹網様体から起こる線維の中にも内側縦束に加わって脊髄へ下行するものがある。第三の線維群としては、動眼神経核、滑車神経核、外転神経核を相互に連絡するものや、顔面神経核や疑核に達するものがある(内側縦束は咀嚼や燕下の協調にとっても重要である)。 D.前庭神経の機能前庭器官は空間定位、眼球の反射運動、平行一般に関与しており、水平面および垂直面に対する頭部の定位にさいしてきわめて重要な役割を果たしている。平衡を保つためには体幹の正中面を垂直に保つ必要があるから頭部の垂直面に対する定位は重要である。また、水平面と視線の方向との関係よりみて、頭部の水平面に対する位置も重要である。 迷路反応と頚の金とは密接な共闘関係にあるが、これは内側前庭脊髄路を介して行われる。このようにして頭部はその正常の位置に保たれるのである。迷路立ち直り反射とは区別しなければならない。迷路反射は卵形嚢と球形嚢からのインパルスによって起こり、重力に対する頭部の位置に関わる。頚部立ち直り反射は頚部の金と頚椎の関節や靱帯にある固有受容器の刺激によって起こる。ネコを高所から落とすと、最初に頚部を正常の位置にまわし(迷路立ち直り反射)、次いで体幹の位置を正にして(頚部立ち直り反射)足から着地する。 E.眼球の反射運動ある特定の対象を凝視し続ける場合には、頭部がどのように動いても眼球はその対象に向かい続ける。このことは向かい合って頭を横に振っている人をみると確かめることができる。その人の眼球は頭の運動と逆に動くのである。頭部の水平面で左側へ回すと、水平半規管の中で無いリンパの流れが起こり、右の膨大部稜では発火頻度がマスのに対し、左の膨大部稜では減少する。膨大部稜からのインパルスは前庭神経核、とくに上核と内側核に伝達され、それらの核のニューロンの中に興奮するものが出来る。これらのインパルスは内側縦束によって眼筋を支配する神経核へと伝達され、右側では外側直筋(外転神経の支配を受ける)が収縮し、内側直筋(動眼神経の支配を受ける)が置換する。左側では外側直筋が弛緩し、内側直筋が収縮する。 前庭神経核は小脳と密接に協調して平衡と姿勢の調節機序に関与する。遠心性線維は内側および外側前庭脊髄路であり、後者は伸筋の緊張を促進する。このことは、上丘と下丘のあいだで実験的に脳幹を切断することによって示すことができる。すなわち、実験動物の四肢の伸筋(抗重力筋)は硬直する(除脳固縮)。前庭神経外側核を選択的に破壊すると除脳固縮はほとんど即座に消失する。
3.蝸牛神経
A.音受容器の構造音受容器は聴覚器の一部であって、迷路の特殊な部位、すなわち、蝸牛管内に存在する。蝸牛管の部分は骨迷路の骨ラセン板上にあり、骨ラセン板の自由縁からは基底板がのびている。基底板は蝸牛の相対する壁にラセン靱帯によって付着している。骨性の蝸牛内の空間は上方の前庭階と下方の鼓室階に分けられている。膜性の蝸牛管は前庭階の下方の部分を占めている。 「蝸牛管と前庭階の境界には前庭膜(Reisner膜)がある。」 前庭階は前庭から始まって、2巻き半して蝸牛頂に達する。前庭階は蝸牛頂で蝸牛孔によって鼓室階に続き、鼓室階は下方で正円窓は第2鼓膜を隔てて鼓室に面している。
B.基底板この膜は内包の骨ラセン板の縁と外包のラセン靱帯のあいだにまたがっている。Corti器官はこの基底板上にある。基底板には内側部(蓋部)と外側部(櫛状部)が区別される。前者は骨ラセン板の縁から蓋膜の自由縁までであり、Corti器官をのせる。後者はラセン靱帯に付着している。 蓋膜はケラチン化した放射状に走る線維から成り、その下面に接して結合組織正の薄い膜がもう1枚ある。蓋膜の鼓室階に面する表面は一層の結合組織性の疎な細胞集団でおおわれている。基底板の幅は蝸牛頂に近いところでは0.5mm、蝸牛底に近いところでは0.04mmで、長さは34mmである。
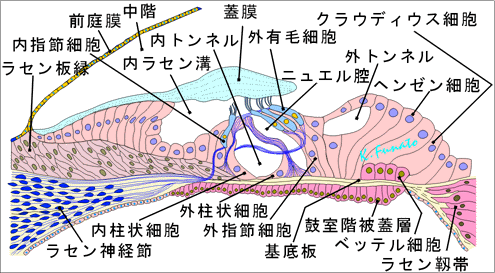
C. Corti器官Corti器官として知られる特殊な感覚上皮は基底板上にある。内ラセン溝の方から純に、次のような要素が区別できる。柱細胞、内有毛細胞、外有毛細胞、Deiters細胞、Hensen細胞、Claudius細胞、網状板、蓋膜。
①柱細胞柱細胞は内・外の2列に並んでいる。これら内柱細胞と外柱細胞は一緒になって一種のアーチを形成しており、このアーチは三次元的には蝸牛の全長にわたって走るトンネルを作っている。内柱細胞の中央部は薄く扁平になっているが、底部は著明に広くなって基底板の蓋部(内側部)上にのっている。また、内柱細胞の頂部には小さな凹みが一つあり、ここに外柱細胞がはまっている。外柱細胞の中央部は円柱状で、両端は広く成っている。内柱細胞の頭板はそれぞれ相対する外柱細胞の頭板をおおうように重なっている。
②蝸牛の感覚細胞蝸牛の感覚細胞は有毛細胞であり、その自由面には多数の不動毛(変形した一種の微絨毛)が出ている。 「この不動毛は聴毛と呼ばれ、長さ3~4μmである。」 不動毛の末端は蓋膜に接している。Cortiのトンネルより外側にある有毛細胞を外有毛細胞、内側にあるものを内有毛細胞という。 内有毛細胞(約3,500)は内柱細胞に相対して一列に並んでいる。これらの内遊毛細胞の自由端は網状膜に取り囲まれており、網状膜は柱細胞の頭板に続いている。内有毛細胞は西洋梨形で、その尖端には50~60本の微絨毛がある。運動毛はないが、細胞面の近縁に近く基底小体が一つある。 「運動毛が退化して基底小体だけが残ったと考えられる。」 不動毛の底部は細くなっていて、電子顕微鏡で見ると、多数の微細線維が縦走し、有毛細胞内につきささって不動毛をささえている。内有毛細胞を上方からみると、不動毛は必ず馬蹄形にならんでいる。 外有毛細胞(約12,000)は外柱細胞の外側に3~4列をなして並んでいる。これらの細胞の列はDeiters細胞(外指節細胞または外支持細胞)の突起によって隔てられている。外遊毛細胞は内有毛細胞よりもずっと長く、その自由端は扁平で、おのおの約100本の不動毛をもつ。上方からみると、不動毛はW形に配列している。不動毛の末端は蓋膜に接している。 「内有毛細胞毛も外有毛細胞もその基底部で支持細胞の上にのっており、基底板までは達しない。」
③Deiters細胞(外支持細胞、外指節細胞)Deiters細胞は紡錘形で、その中央部は太く、両端は細く成っている。底部の突起は基底板にのっているが、先端部の突起は細く伸びて有毛細胞の自由端にまで達し、そこで指節と呼ばれる指状の拡がりを作って終わっている。指節は蓋膜を取り除いてCorti器官を上方からみるととくによくみえる。 「拡がった指節は互いに密接に網状膜を作り、この毛状膜を貫くようにして外遊毛細胞が突出している。Deiters細胞はこのようにしてそれぞれ1個の外遊毛細胞を支えている。」 d)内支持細胞(内指節細胞)は内柱細胞の内側に1列に並ぶ細胞で、内有毛細胞の支持細胞である。Deiters細胞と同様、基底部は基底板にのり、先端部は細く伸びて有毛細胞の自由端に達し、指節を作って拡がって終わる。
④蓋膜この膜はCorti器官の上をおおう屋根とみることができる。その内側縁はラセン板縁に付着する。蓋膜は外層部ではずっと厚くなっており、外遊毛細胞の上方で鋭い自由縁をもって終わっている。蓋膜はゼリー状の屈折率の高い物質で、内部にはきわめて微細な線維がネットワークを作って埋まっている。
⑤神経終末の構造内有毛細胞の基底側には蝸牛神経の終末が接触する。これらの終末には求心性線維のものと遠心性線維のものとがある。蝸牛神経を中枢へと向かう求心性線維の終末には多数の電子密度の高い封入体や層状体などがみられる。中枢から起こる遠心性線維の終末にはミトコンドリア、微小管、シナプス小胞がみられるが、これら終末は内有毛細胞には結合せず、求心性線維の終末とシナプス結合する。 「内有毛細胞の細胞体とシナプス結合する遠心性線維終末の存在も報告されている。」 蝸牛神経線維約30,000本のうち、約18,000本が内有毛細胞と接している。すなわち、約3,500の内有毛細胞からの情報が約18,000本の蝸牛神経線維へ拡散する。 外有毛細胞の神経支配のパターンは内有毛細胞のそれとは異なる。すなわち、すくなくとも12,000の外有毛細胞が約4,000本の蝸牛神経線維に接触する。外有毛細胞の細胞体はその大部分がDeiters細胞でおおわれており、蝸牛神経線維とシナプス結合する部位だけがDeiters細胞でおおわれていない。中枢から起こる遠心性線維は、内有毛細胞の場合とは違って、外有毛細胞の細胞体と直接シナプス結合する。
D.蝸牛の機能鼓膜の振動は耳小骨によって卵円窓(前庭窓)に伝達される。アブミ骨底の動きによって外リンパが振動するから、空気の振動(音)が液体の振動に変換されることになる。外リンパは圧縮されないからアブミ骨の内方への動きは正円窓(蝸牛窓)にある第二鼓膜の外側方への動き、すなわち鼓室へ向かっての動きによって代償される。外リンパの振動は振幅は小さいが、圧力の変化は大きい。前庭階と鼓室階の外リンパの振動によって蝸牛管も振動する。すなわち、基底板がその表面と垂直面で動き始める。 蓋板と基底板の相対運動によって加わる剪力ののために聴毛が曲げられ、これが刺激となって聴神経活動電位が発生する。活動電位の発生する部位は基底板の移動最大の部位に一致しており、高音は蝸牛管の底部の受容細胞を興奮させるのに対して、低音は蝸牛管の先端部の受容細胞を興奮させる。 「蝸牛底にある基底板は短く、蝸牛頂にある基底板は長い。したがって、基底板の振動が最大になる領域は低音では蝸牛頂に近く、高音では蝸牛底に近い。」
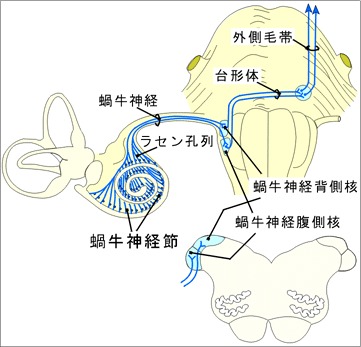
E.蝸牛神経線維蝸牛神経線維はラセン神経節の双極細胞の中枢性突起である。これらの線維は脳幹にはいるとただちに上行枝と下行しに分かれ、前者は腹側蝸牛神経核、後者は背側蝸牛神経核でシナプス結合する。
4.聴覚の中枢神経路聴覚機能に関与するニューロン回路は菱脳、中脳、間脳、大脳皮質の中枢が複雑に連絡しあって構成されている。これらの入力性感覚神経路に加えてさらに遠心性の(上)オリーブ(核)蝸牛束がCorti器官の有毛細胞の興奮性を抑制的に調節している。
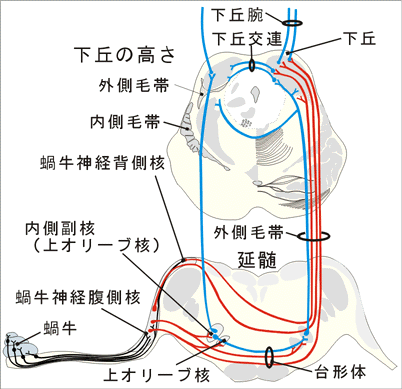
A.菱脳の聴覚系背側蝸牛神経核は第四脳室底の最外側部に隆まりをなしている。多くの哺乳類でこの核には層構造が認められるが、「人では認めがたい。」腹側蝸牛神経核は中等大の細胞より成る。この悪には局所的な構造の差異があるが、層構造はみられない。蝸牛の底部からの線維(高音を伝達する線維)は背側及び腹側蝸牛神経核の背側部でシナプス結合し、一方、蝸牛の先端部からの線維(低音を伝達する線維)は両蝸牛神経核の腹側部でシナプス結合する。蝸牛神経核から出る線維は両側性に菱脳の次の核に投射する。
1)背側蝸牛神経核からの線維は下小脳脚の背側方を通って脳幹網様体を横切り、正中線で交叉して外側毛帯に加わる。脳幹網様体を横切る線維は2群の線維束をなしている。背側聴条(Monakow束)と中間聴条(Held束)である。聴条の線維の多くが上オリーブ核と台形体核でシナプス結合する。 「ただし、背側聴条の線維は直接下丘に達する。」
2)腹側蝸牛神経核からの線維は腹側聴条で、被蓋腹側部を横走し(台形体)、正中線で交叉する。これらの線維の一部は脳幹網様体、台形体核、上オリーブ核のレベルで鋭く上方に屈曲して、いわゆる外側毛帯となって上行する。 上オリーブ核は内側毛帯と三叉神経脊髄路核とのあいだに存在する核群であり、両側の蝸牛神経核から求心性線維を受け、遠心性線維は外側毛帯に加わる。 台形体核は台形体の線維のあいだに散在する細胞より成り、台形体の線維や聴条の線維がこれとシナプス結合する。 「上オリーブ核群には、台形体腹側核、台形体背側核、内側上オリーブ核、外側上オリーブ核、腹側上オリーブ核、背側上オリーブ核が区別されるが、ヒトでは一般に小さく、腹側上オリーブ核や背側上オリーブ核は指摘できない。また、台形体核もほとんどみえない。」 外側毛帯:蝸牛神経核や上オリーブ核から出て、被蓋の外側部を上行して下丘に向かう交叉性および非交叉性の線維群である。これらの線維のあいだにはニューロンが散在している(腹側および背側外側毛帯核)。外側毛帯の船員中にはkろえらのニューロンとシナプス結合するものがある。外側毛帯は脳幹の聴覚路の中でもっとも重要なものである。
B.中脳の聴覚系中脳の聴覚路の主要な核は下丘である。下丘には種々の型の細胞を含む中心部と、4層の細胞層が認められる皮質部が区別される。 下丘への求心性線維にはつぎのようなものがある。
1)外側毛帯の線維は中心部と皮質部の深部の二層でシナプス結合する。
2)反対側の下丘からの線維(下丘交連)。
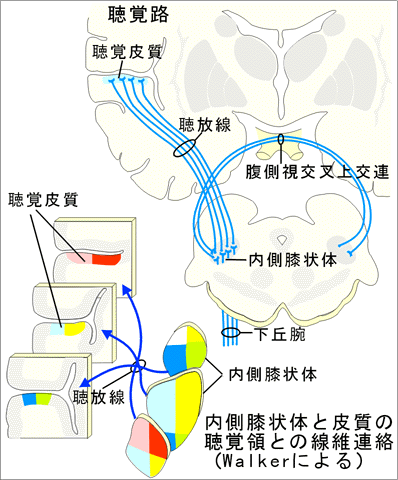
3)大脳皮質からの線維は皮質部の浅層でシナプス結合する。 下丘からの遠心性線維は下丘腕を通って視床の内側膝状体へ向かう。その他の線維には反対側の下丘へ向かうものや、菱脳の聴覚系神経核へと下行するものなどがある。
C.間脳の聴覚系下丘からの線維は内側膝状体でシナプス結合する。内側膝状体には小細胞部と大細胞部が区別され、下丘からの線維は小細胞部でシナプス結合する。このさい、音階対応投射がみられ、低音は外側部に高音は内側部に投射する。
D.聴覚系の大脳皮質への投射内側膝状体ニューロンは内側膝状体側頭路を通じて横側頭回(41野、42野)に軸索を送る。動物においては音階対応配列が聴覚領皮質で証明されている。すなわち、蝸牛の底部の高音受容部は41野の前部に投射し、蝸牛の先端部の低音受容部は41野の後部へ投射する。一側の蝸牛が両側の聴覚領皮質に投射する。 「Pfeifer(1936)によれば、ヒトの一次聴覚領に音階対応配列があるとすれば、高音受容部は内側部に低音受容部は外側部に位置するはずであるという。」 上記のように聴覚系では多くの中継核が連結している。これらのうち、上オリーブ核、下丘、内側膝状体などは主として湯シ列狽ノ連結されているのに対し、台形体核、脳幹網様体、外側毛帯核などは燈タ列狽ノ連結されている。これらの中継核が単なる湯・p点蝿ネ上の働きをしていることは明らかであり、音色や音の強さの識別、有意味音と雑音の識別など、いわゆる側方抑制の騎乗がかかわる聴覚機能はこれらの中継核によって左右される。
E.聴覚神経路内の下行性線維中枢神経系は聴覚上方の入力を調節している。このような機能にかかわる下行性悼盗S柏・ロはすべての聴覚系の核から起こり、下位の核でシナプス結合する。これらの下行性線維の中でもっとも有名なのが(上)オリーブ(核)蝸牛束である。この線維束は上オリーブ核群「またはその近傍」から起こり、同側性および反対側性要素から成る。上オリーブ核蝸牛束は約500本の線維を含み、前庭神経に沿って脳幹を去り、迷路の中で前蝸牛吻合を通って前庭神経から離れて蝸牛神経に加わる。これらの線維は主として外有毛線維の細胞体の底部および内有毛細胞と連絡している求心性線維終末とシナプス結合する。 「上オリーブ核蝸牛束以外の下行性繊維として、聴覚領から同側の内側膝状体や下丘にいたるもの、下丘から主として同側の上オリーブ核群へいたるもの、外側毛帯核背側核から両側の上オリーブ核群へいたるもの、外側毛帯核腹側核から反対側の蝸牛神経腹側核にいたるもの、などが報告されている。また、聴覚系では交連線維が発達しており、左右の聴覚領皮質を結ぶ脳梁線維の他、下丘交連、上下の外側毛帯背側核間を走るProbst交連、左右の上オリーブ核群間を走る台形体や聴条がある。」
5.内耳神経の臨床的側面前庭神経及び蝸牛神経は、本質的に独立しているが、両者が接近しているために、末梢の損傷によりしばしば臨床症状が併発する。そのため内耳神経の損傷は耳鳴り、めまい、平衡感覚の損傷、聴覚の障害や消失をおこす。内耳神経の中枢損傷は、蝸牛神経と前庭神経のどちらかを巻き込む。難聴(以下、聴覚障害とする)には2種類ある。 ・伝播性聴覚障害は、外耳ないし中耳疾患(たとえば中耳炎により起こる)。 ・感音性聴覚障害は、蝸牛ないし蝸牛から脳への経路の疾患により起こる。 内耳神経腫は、成長の遅い良性のSchwann細胞の腫瘍で、前庭神経が内耳道を通る当たりで起こる。その初期症状は聴覚の消失である。 左または右の聴覚領大脳皮質が左右の蝸牛神経核から発する信号を受ける(べつの言い方を、右の蝸牛神経核より発する信号も左の蝸牛神経核より発する信号も、それぞれが両側の皮質聴覚領に入力される)という点である。聴覚系にみられるこのような両側性伝導の臨床的意義は明白であり、たとえば右の大脳皮質で聴覚領に損傷が生じた際でも、患者は左右の耳からの聴覚刺激を健常な左の大脳半球で認識することが出来るであろう。このことは、単に右の皮質聴覚領の損傷時ばかりでなく、右の内側膝状体、下丘核、外側毛帯など聴覚伝導路全体の一側性損傷の場合にもあてはまる。しかし右の内耳神経に含まれている第一次ニューロンが、内耳から蝸牛神経核までの経路のどこかで切断されたり、損傷した場合には、右耳の聴力が失われる。また、左の同様病変は左耳の聴力消失をもたらす。下丘核より出る線維の中には、種々の下位運動ニューロンに達するものがあり、これが聴覚反射運動(大きな音に急にさらされたときの眼を閉じる、身体が跳ね上がる、などの動作がその一例をもたらす。 米国では聴覚消失に悩む人の数が数百万にも達している。この状態は2種類に区別しうるが、その第一は伝音性聴覚消失、すなわち内耳の伝音系の故障のために蝸牛に聴覚刺激が伝わらないケースである。この場合の故障には鼓膜破裂、耳管閉鎖なども該当しうる。しかし伝音性聴覚消失の原因疾患として最も高頻度に認められるのは、耳硬化症(中耳のアブミ骨が可動性を失い、そのため蝸牛への伝音がたたれることになる)である。第二の種類をなすものが感覚神経性聴覚消失であり、これは名が示すように蝸牛または内耳神経の損傷によりもたらされる。 聴覚障害が伝音性のものか感覚神経性のものかを識別することは、診断、治療のうえで重要である。伝音性障害では発振音叉が耳元に存在するだけでは聴覚が生じないが、その音叉を頭蓋骨に当てれば聴覚を生じる(中耳を経由しない骨伝導経路で振動が蝸牛に達するためい。一方、感覚神経性障害では発振音叉が耳もとに置かれたときも、頭蓋骨に当てられているときも、聴覚を生じない。 近年、伝音性聴覚障害(ことに耳硬化症によるもの)の治療法が大きく進歩している。それは顕微鏡手術により固着しているアブミ骨を可動化したり除去するなどの方法を主体とするものであって、患者の聴覚に大きな改善をもたらす。感覚神経性聴覚障害の原因は実に多様であるが、その主なもののみを以下に記す。
1.妊娠の風疹感染(新生児の聴覚完全障失を、非常にしばしばもたらす)
2.ある種類の抗生物質(ストレプトマイシン、ゲンタマイシン、ナオマイシン)の大量投与。これは、しばしば前庭機能機能失調をも伴うような、部分的あるいは完全な聴力消失を来しうる。
3.蝸牛の萎縮。高齢者における聴力消失例の多くが、この原因による。
4.聴神経鞘腫
5.遺伝子異常(種々のタイプの先天性聴覚消失をもたらす) 聴力を以前には備えていたが今は感覚性聴覚消失の状態に陥っている患者のために、「バイオニックス耳」ともいえる装置が、光学と医学の研究者たちの協力でつくられている。集音用マイクロフォンと小型アンプを患者は装着し、そのアンプから出力を伝える特殊電線の終端が患者の蝸牛に埋め込められというものであり、結果は非常によい。しかし先天性の聴力障害、幼児期に発症した感覚神経性聴覚消失に対しては、この装置も効果を示さない。
解剖学用語(内耳神経)1. 内耳神経 [VIII] ラ:Nervus vestibulocochlearis [VIII] 英:Vestibulocochlear nerve [VIII] →第VIII脳神経。内耳神経は内耳に分布する知覚神経で、その終止核は延髄および橋背部にある。2根、すなわち上根(前庭根)と下根(蝸牛根)とをもって、橋の下縁で顔面神経の外側において脳を去り、合して半月状の断面をなして顔面神経を外側から包んで内耳道に入り、ここで上根は前庭神経に、下根は蝸牛神経に移行する。
2. 前庭神経 ラ:Nervus vestibularis 英:Vestibular nerve →前庭核より平衡斑および膨大部稜へいたる第VIII脳神経の部分。
3. 前庭神経節 ラ:Ganglion vestibulare 英:Vestibular ganglion →内耳道床中にあり、双極細胞よりなる神経節。
4. 蝸牛交通枝 ラ:R. communicans cochlearis 英:Cochlear communicating branch
5. 上部 ラ:Pars superior 英:Superior part →前庭神経節上部。前および外側半規管、卵形嚢、および球形嚢の前部へ分布する。
6. 卵形嚢膨大部神経 ラ:Nervus utriculoampullaris 英:Utriculo-ampullar nerve →前庭神経の上枝。内耳道底の上前庭野を通り、さらに上篩斑を経て前庭に入り卵形嚢神経、前膨大部神経、外側膨大部神経の3枝に分かれる。
7. 卵形嚢神経 ラ:Nervus utricularis 英:Utricular nerve →卵形嚢斑の感覚上皮にいく。
8. 前膨大部神経 ラ:Nervus ampullaris anterior 英:Anterior ampullary nerve →前半規管の前膨大部稜の感覚上皮に行く。
9. 外側膨大部神経 ラ:Nervus ampullaris lateralis 英:Lateral ampullar nerve →外側半規管の外側膨大部稜の感覚上皮に行く。
10. 下部 ラ:Pars inferior 英:Inferior part →球形嚢神経と後膨大部神経とがあり、球形嚢神経は球形嚢斑の感覚上皮に分布し、後膨大部神経は内耳道において最も早く神経管から分かれて、内耳道底の単孔を通りさらに下篩斑を経て後膨大稜の感覚上皮に終わる。球形嚢神経は発生学的には蝸牛神経と特に関係が深いので、蝸牛神経として取り扱う人もあるが、その神経節は前庭神経節である。
11. 後膨大部神経 ラ:Nervus ampullaris posterior 英:Posterior ampullary nerve →後半規管の膨大部への感覚上皮に終わる枝。
12. 球形嚢神経 ラ:Nervus saccularis 英:Saccular nerve →球形嚢斑の球形嚢斑の感覚上皮に終わる枝。
13. 蝸牛神経 ラ:Nervus cochlearis 英:Cochlear nerve →蝸牛神経は内耳道底でラセン孔列を通り、蝸牛軸縦管に入り、次第に蝸牛軸ラセン管には入り、そのなかにあるラセン神経節を経てラセン器の感覚上皮に至る。
14. 蝸牛神経節;ラセン神経節 ラ:Ganglion cochleare; Ganglion spirale cochleae 英:Cochlear ganglion; Spiral ganglion →蝸牛軸のラセン板底部に沿って存在するラセン状の神経細胞索。
|