Spalteholz HANDATLAS DER ANATOMIE DES MENSCHEN VON WERNER SPALTEHOLZ
メニューは解剖学(TA)にリンクしてあります。図の番号をクリックすると下記の説明へ、右側の用語をクリックすると解剖学(TA)にジャンプします。
831


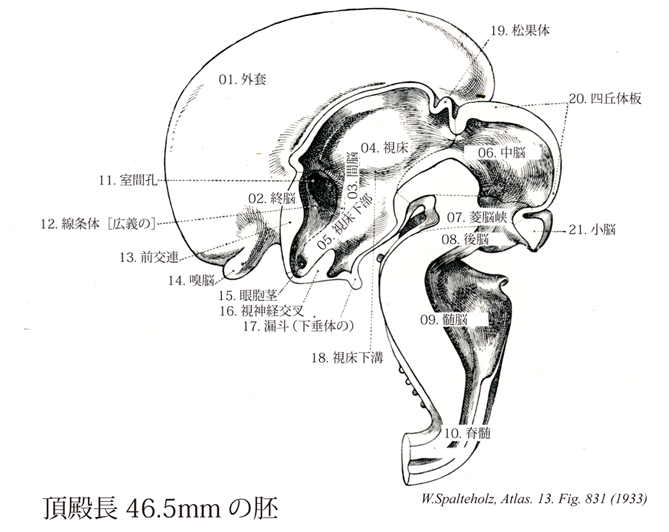
- 831_01【Pallium外套;大脳皮質 Pallium】
→(大脳半球のうち脳幹を取り囲む部分。)
- 831_02【Telencephalon終脳 Telencephalon】 Part consisting of two hemispheres that each surround a lateral ventricle and are connected with each other.
→(「末端」を意味するギリシャ語のtelosとencephlonを結合した言葉である。Encephalonは、「~の中に」という意味のギリシャ語の接頭詞enと、「頭」を意味するギリシャ語であるkephaleを一緒にしてつくられたもので、「頭の中にあるもの」、すなわち「脳」を指す。Telencephalonは、脳の末端部という意味である。 終脳は最高次機能の中枢であり、人脳で著しい発達を示す。大脳半球と基底核を合わせたものが終脳であるが、後者は粗大運動性の領域で大脳皮質に覆われた位置を占める(脳を切断しないと観察できない)。左右の大脳半球はこれに対し非常に大きな構造で、大脳縦裂を相互の境界とし、かつ外見上での脳の大部分を占めている。大脳半球の外表面には大脳回と呼ばれる曲がりくねった高まり、大脳回1つ1つの境をなす大脳溝と呼ばれる浅い溝(深いものは・・・裂と呼ぶ)が認められる。大脳回・溝の多くは人脳に共通するが、しかしそれのまったく同じ配列パターンがみられることは、たとえ同一個体の左右の大脳半球を比較した場合でも、また他人同士の脳を比較した場合でも、決してない。中心溝と大脳外側溝を使い大脳半球を4領域(前頭葉:中心溝より前、頭頂葉:中心溝より後、頭頂後頭溝と大脳外側溝をつなぐ仮想腺まで、側頭葉:大脳外側溝より下、後頭葉:頭頂後頭溝と大脳外側溝をつなぐ仮想線より後)に分けることができる。これら4区分のそれぞれ、固有の特殊中枢がある。たとえば前頭葉の中心前回(中心溝のすぐ前に位置)は随意運動司令センターであり、前頭葉の前端部、すなわち前頭極は人格の座(これの損傷が人格変化を招く)とされる。脳の下面(基底面に同じ)でも終脳の広がりが見られる。すなわち、複数の眼窩回やそれらに接近する嗅神経(味覚を伝える神経)などに注意されたい。左右の視神経は互いに近づき視神経交叉を示した後に、再び左右の視索に分かれ後方へ向かう。側頭葉の一部である海馬傍回と、これの特徴的突出部分をなす鈎も脳の基底面で認められる。脳の水平断面では左右の大脳半球が表層の灰白質すなわち皮質(神経細胞体が集まる部分)と深層の白質(有髄神経線維の密集部分)からなることがわかる。左右の半球皮質をつなぐ神経線維を交連線維と呼ぶが、これの大集合体が脳梁にほかならない。同一半球内で葉間、あるいは脳回間をつなぐ長短様々の神経線維が連合線維であり、大脳半球皮質とそれ胃が引中枢神経部分とをつなぐ(皮質より下行、あるいは皮質に向かい上行する)神経線維が投射線維である。投射線維の集合体、すなわち内包を前脚・膝(中間部に当たる部分)・後脚に区分するが、膝のすぐ外側に基底核の一部(淡蒼球・前障など)が位置している。大脳半球表面について前述の4区画(葉)のほかに、さらに1~2の区画を設けることがある。大脳外側溝を押し広げると、その奥に島と呼ばれる高まりが現れるが、これを第5番目の葉としてよい。人脳の島がどのような機能を営むかについては、不明な点が多い。島を覆う位置にある、前頭・頭頂・側頭の各葉の大部分(大脳外側溝沿い)は弁蓋と呼ばれる。第6番目の葉は辺縁系に属するもので、これには帯状回、海馬傍回、歯状回なが含まれる。 )
- 831_03【Diencephalon間脳 Diencephalon】 Part of the forebrain comprising the thalamus, epithalamus and pineal body, hypothalamus, and globus pallidus. It encloses the third ventricle and extends from the anterior border of the superior colliculus as far as the interventricular foramen.
→(間脳は「間」を意味するギリシャ語の接頭詞diaと、「脳」を意味するencephalonを結合したもの。間脳は中脳の前方で第三脳室を取り囲んだ領域をいう。背側方は側脳室におおわれ、背外側は分界条によって尾状核と境され、外側を内方によって取り囲まれている。前方は室間孔まで伸び、後方は後交連と乳頭体の後方を結ぶ線で中脳被蓋に移行する。間脳はさらに背側視床、視床下部、腹側視床および視床上部に分かれる。背側視床はこれらのうちもっとも大きな部位を占め左右を視床間橋(中間質)によって結ばれる。背側視床と視床上部とを視床脳とよぶことがある。 間脳の発生development of the diencephalon:間脳は間脳胞から発生する。胎生第4週のおわりころにおける脳の原基は、前脳胞・中脳胞・菱脳胞の3脳胞が確立し、前脳胞ではその外側壁の前腹側部から外方に向かって大きい眼胞が膨出している。第5週に入ると(第14段階)、この眼胞の出発部の前背側にあたる部分の外側壁が前外方、ついで背外方に向かって膨出し、左右1対の半球胞を形成する。左右の半球胞を連ねる前脳胞の前背側端部を終脳正中部という。終脳正中部と半球胞とが終脳を形成し、これまでの前脳胞の大部分を占める領域は、これ以降、間脳胞と呼ばれる。 間脳胞ははじめ頭尾(前後)方向に長い管であり、その内腔は間脳室(後に第三脳室)とよばれる。陥凹胞においても、中脳以下の神経管の各部におけると同じく、実質的な神経細胞の形成は左右の外側壁においてのみおこり、背側壁(蓋板)と腹側壁(底板)においては著明な肥厚は見られない。蓋板は単層立方上皮となり、外側から間葉組織によって裏打ちされて、第三脳室脈絡組織となる。 肥厚・増大していく間脳の外側壁の内面には一過性に前後に走る3本の浅い溝(背側から腹側へ1.間脳背側溝、2.間脳中間溝、3.間脳腹側溝)がみとめられ、これによって外側壁は1.視床上部、2.背側視床、3.腹側視床、4.視床下部の4部に区画される。その後の発育において背側視床と腹側視床がとくに高度に発育し、両者が合一して強大な視床を形成する。胚芽層から生じた多数の神経[芽]細胞は外套層および縁帯の各所に集合して前核・内側核・中心核・腹側核・外側核・視床枕核などの視床核を形成する。 視床上部はヒトでははなはだ退化的で、これに属する構造物としては、間脳の後端部における松果体、手綱および手綱三角のみである。松果体は胎生第7週において間脳の蓋板の後端正中部から背後方に向かって生ずる1個の中空の膨出として発生する。 視床下部は間脳の外側壁および腹側壁を埋め、およそ8個に大別される神経核を形成する。 間脳胞の内腔である間脳室は、はじめは円形に近い横断面を示すが、外側壁の発育につれて上下方向に広く左右方向に狭い空間となり、ついには正中矢状断面に一致した左右の幅の非常に狭い裂隙状の腔となる。この間脳室とその前端につづくごく狭い終脳正中部の内腔を合わせて第三脳室という。)
- 831_04【Thalamus; Dorsal thalamus視床;背側視床 Thalamus】 Part extending from the interventricular foramen to the tectal plate. Medially it borders on the third ventricle, laterally on the internal capsule and basal ganglia. It is formed by a collection of nuclei derived during ontogenetic development from the dorsal thalamus.
→(視床は、間脳の大きいほうの背側部分を形成する灰白質。背側間脳溝と視床下溝の間の領域であるが、発生の間に大きく発育して、間脳背側部の広い範囲を占めるようになる。間脳は個体発生上、背側視床、腹側視床、視床下部および視床上部の四つの部位に分けられるが、その中で最も大きな部位を占めるのが背側視床である。単に視床といった場合は背側視床を指す。視床は第三脳室の両壁をなす卵円形の構造で、背側の遊離面は薄い髄質から成る帯層におおわれ、肺内側端に視床上部の構造である視床髄条が、前端より後方に走り手網核に付く。また背外側端は分界条によって終脳の尾状核と、外側方は外髄板によっておおわれ腹側視床の視床網様核と境されている。左右の視床は第三脳室内にまたがる視床間橋(中間質)によってつながり、視床下溝で視床下部と境される。視床の内部を構成している視床核は視床脚を介して大脳皮質と相互に結合する。内部には内髄板とよばれる線維板視床を内側部、外側部および前部に分けている。視床は感覚系と統合系との非常に重要な連絡部位である。嗅覚路以外のすべての感覚路がそれぞれ相当する視床の領域に投射する。「最近の研究によれば、嗅覚系も視床を投射する可能性がある」。視床で処理された感覚系情報の流れは視床大脳皮質線維を経て大脳皮質へと送られるが、大脳皮質の側からは多数の大脳皮質視床線維を介して視床における情報処理系に影響が及んでおり、したがって、視床と大脳皮質とは一つの機能単位としてはたらく。「運動」情報は小脳と大脳基底核を経て伝達され、統合系(大脳辺縁系や脳幹網様体など)からのさらに複雑な情報も視床に達する。したがって、視床は一方では大脳辺縁系と脳幹網様体との連結点として機能し、他方では大脳皮質も連絡しているわけである。)
- 831_05【Hypothalamus視床下部 Hypothalamus】 Basal portion of the diencephalon. It consists of the floor and, from the hypothalamic sulcus onward, the basal portions of the walls of the third ventricle. It extends from the lamina terminalis to just posterior to the mammillary body and is visible on the base of the brain. It is composed of individual areas that can be distinguished histologically to a greater or lesser extent. It functions as an autonomous control center sending hormonal and nerve efferents.
→(視床下部は間脳の中で、内臓機能、自律機能および内分泌機能と最も関係が深い部分である。これらの機能のすべてが、感情的および情動的行動と密接に関係している。第三脳室の側壁の下部および底にあたる。脳底面からみると、吻側から数えて、視交叉、漏斗、灰白隆起、乳頭体とつづき、漏斗の先端は下垂体に連なる。背側は視床下溝により視床と境されており、吻側は終脳の視索前野に、尾側は中脳被蓋と中脳中心灰白質に、尾外側は視床腹側部に移行する。通常、矢状面に平行な三つの帯状領域、すなわち、視床下部脳室周囲層、視床下部内側野、視床下部外側野に区分される。これらの間を多数の細かい神経線維が主として吻尾方向に走っている。視床下部は前後径が約10mmである。視床下部は内部環境を正常に維持する機序に関与し、また心悸亢進、瞳孔散大、“冷汗”の分泌などの情動反応の表出にも一役を演じている。大脳皮質を除去し、背側視床を除去した後でも、怒り反応はあらわれる(「みかけの怒りSham rage」)。さらに、視床下部は成長、性的成熟など新地あの成熟過程にも関与している。したがって、視床下部を損傷すると、広汎で顕著な内分泌性、代謝性、行動性(情動性)の以上が一緒に起こってくることになる。視床下部は下垂体ホルモンによって内部環境に影響を及ぼすが、さらに脳幹網様体や自律神経系を介しても同様の働きを示す。視床下部への情報は通常の入力神経系によって伝達されるばかりでなく、視床下部のニューロンは内部環境からの物理的および化学的刺激(血液の温度など)にも反応する。植物性機能の中には、その統御中枢が視床下部自体に存在するものもあるが、呼吸や心臓の活動などの機能に関しては、視床下部はもっと下位の中枢に対する修飾器として働いている。)
- 831_06【Mesencephalon; Midbrain中脳 Mesencephalon】 Part of the brain that does not form a genetic unit, but rather developed in the region between the rhombencephalon and prosencephalon. This topographical term also includes the tegmentum, red nucleus, tectal plate, cerebral crura, and substantia nigra.
→(中脳は「中央」を意味するギリシャ語の接頭詞mesoと、「脳」を意味するencephalonを結合したもの。中脳、橋、延髄を合わせて脳幹と呼ぶが、これは頭蓋底の大後頭孔の所から上方に向かい、大脳半球の基底面にまで伸びる楔形の構造である。中脳は狭義の脳幹の最上方部で、上方に間脳、下方に橋との間の中脳水道を囲む比較的上下に短い構造を指す。間脳との境は厳密には不明確であるが、背側に後交連の後部、腹側に乳頭体の後方を通る面で境される。下方は背側に下丘の後方と腹側の橋の前方を通る面で比較的明確に境される。外形を見ると背側に蓋板によって形成された4個の隆起があり上方の一対を上丘、下方の一対を下丘という。上丘および下丘からは上外側に線維束を出し、それぞれ上丘腕および下丘腕として間脳につづく。腹側には大脳脚がみられ、その間に多数の小血管が通る後有孔質の間の細い溝を大脳脚内側溝とよび、ここから動眼神経の根がでる。断面では背側部は蓋板で包まれ、(視蓋とも呼ばれる)上丘および下丘を形成し、その腹側端はほぼ中脳水道の中央部を通る面で区切られる。これより腹側を広義の大脳脚というが、これはさらに中脳被蓋と狭義の大脳脚にわけられる。中脳では固有の細胞集団と線維束があり、細胞群としては中脳水道を取り囲む中心灰白質が三叉神経中脳路および核によって外側を包まれ、腹側正中部には上方に動眼神経核、下方に滑車神経核が存在する。また上方の動眼神経核の腹外側に赤核があり、さらに腹側に大脳脚の背側面を覆って黒質が存在する。正中腹側端部の大脳脚にはさまれた部位には脚間核がある。中脳に出入りする線維束で著明なものは中心灰白質内には腹外側部に背側縦束があり、赤核の背側および背外側方に中心被蓋路がある。さらに上丘中央から下丘の高さで正中部に強い線維の交叉がみられる。これらの交叉は被蓋交叉および上小脳脚交叉で、被蓋交叉の背側部は多くは上丘および上小脳脚交叉で、被蓋交叉の背側部は多くは上丘および被蓋からの下行線維から成り、腹側部は赤核からの下行線維から成る。また上小脳脚交叉は小脳核から赤核および視床へ投射する線維の交叉部である。)
- 831_07【Rhombencephalic isthmus菱脳峡 Isthmus rhombencephali】 Genetic, structural, and functional unit. Systematically, it comprises the medulla oblongata, pons, and cerebellum. It surrounds the fourth ventricle.
→(「菱形」を意味するギリシャ語のrhombosと、encephalonを結合したものである。 菱脳は3個の脳胞(前脳、中脳、菱脳)のうちの最尾側のもので、脊髄の頭側につづき頭側半の後脳(橋と小脳)と尾側半の髄脳(=延髄)の2区分される。菱脳の発生において特異なことは、第4週の終わりごろ(第12段階)から蓋板が非常に薄くなるとともに左右にはなはだ広くなるところである。この広く薄くなった蓋板を菱脳蓋という。菱脳蓋の幅は菱脳の中央部(後脳と菱脳の移行部)で最も広く、それより頭側および尾側で次第に狭くなり、菱脳蓋は全体として頭尾方向に細長い菱形となる。頭側の中脳との境界のくびれ(菱脳峡)は第4週の中頃(第11段階)から認められる。蓋板の変化に応じて、はじめ菱脳室の左右の壁をなしていた翼板と基板は、次第に外方に倒れていき、結局、菱脳室の底をつくることになり、全体として菱形窩とよばれる。こうなると底板は菱形窩の正中部を頭尾方向に走る正中溝となり、翼板と基板を境する境界溝は、同名の溝として、正中溝の外側で凸面を外方に向けた弓形をなして頭尾方向に走るようになる。このようにして菱脳室は腹背に扁平で、頭尾に長く、左右に広い菱形の腔となり、第四脳室とよばれる。菱脳蓋は外から間葉組織によって裏打ちされて第四脳室脈絡組織となる。翼板と基板では胚芽層・外套層・縁帯の分化がおこり、外套層は神経細胞で充たされる。このれらの神経細胞は脊髄におけるようなひとつづきの灰白柱をつくらず、いくつかの灰白質塊に断裂する。このような灰白質塊(神経細胞の集団)を神経核という。翼板からは知覚性の、基板からは運動性の脳神経核が生ずるが、これらの配列には整然とした規則性がみられる。基板においては、内側から外側に向かって、①頭部体節由来の骨格筋を支配する体運動核群(M1)、②鰓弓由来の骨格筋を支配する特殊内臓運動核群(M2)、③内臓の平滑筋や腺を支配する一般内臓運動核群(M3)が分化し、翼板においては、同様に①内臓からの求心線維を受け入れる一般内臓知覚核(S1)、②鰓弓領域に発する味覚線維を受け取る特殊内臓知覚核(S2)、③頭顔部の皮膚からの知覚線維を受け取る体知覚核(S3)と④内耳からの求心線維を受ける特殊体知覚核(S4)が分化する。基板および翼板からは、以上の諸核をつくるもののほかに、多数の神経細胞が発生する。これは特別の細胞集団をつくることなく、外套層の中に散在し、これらの神経突起は同側性および交叉性に上行・下行して、脳および脊髄の下腔bに達する。このようにして特定の神経核以外の部分では、外套層は交錯する神経線維の間に神経細胞が散在する状態となり、網様体と名づけられる。また交叉性神経線維はすべて底板の縁帯は交叉線維に満たされて著しく肥厚し、正中縫線となる。翼板と蓋板の移行部を菱脳唇という。後脳の菱脳唇は巨大に発育して小脳を形成する。髄脳は菱脳唇は多数の神経細胞を生ずるが、これらは縁帯の中を腹内方に遊走し、基板の縁帯の中に大きい神経核をつくる。頭側部から生じた神経細胞は後脳の腹側部に橋核、尾側部から生じたものは髄脳の腹側部にオリーブ核を形成する。)
- 831_08【Metencephalon; Pons and cerebellum後脳;橋と小脳 Metencephalon; Pons et cerebellum】 Part of the rhombencephalon consisting of the pons and cerebellum.
→(「後ろ」を意味するギリシャ語の接頭詞とmetaと、「脳」を意味するencephalonを結合して作られた言葉である。 Metencephalonは小脳と橋を意味する。しかし、hindbrainは後脳と延髄、すなわち菱脳の意味で用いられることがおおい。また、Hinterhirnは橋と同義に用いられていることがある。小脳は表面に細かなヒダを多数備えた、後頭葉直下に位置するような脳部分であって、平衡感覚・筋緊張調節・筋活動強調などに関係している。橋は中脳と延髄の間に位置し、第四脳室を隔てて小脳と向き合う環形にある。多種の上行性あるいは構成伝導路が橋を通り抜けるが、第5~7脳神経(三叉神経・外転神経・顔面神経)の諸核も橋内部に存在している。)
- 831_09【Myelencephalon; Medulla oblongata; Bulb髄脳;延髄;脊髄球 Myelencephalon; Medulla oblongata; Bulb】 Rostral continuation of the spinal cord that ends cranially at medullopontine sulcus at the posterior border of the pons. Its agreedupon caudal border lies along a plane above the root fibers of the first cervical segment.
→(脳幹の最下部に位置し、直接脊髄に連続する。錐体交叉の下部境界から上にのびて橋に至る。Mylencephalonは、「脊髄」を意味するギリシャ語のmyelosと、「脳」を意味するencephalonを結合してつくられた言葉で、脳の中で一番脊髄側にある部分を指す。Medulla oblongataという語は、元来脊髄の吻側への延長部、すなわち脳全体を指す言葉であったが、後に脳のなかで脊髄に隣接した領域だけを指すようになった。延髄は、脊髄に比べやや膨らんでいるので、bulus「球」とよばれることがある。 脳幹の最尾部で尾方は第一頚神経の根を境として、脊髄に、吻側は橋に移行する。①外表面:外表面には脊髄の前正中裂、前外側溝、後外側溝および後正中溝につづく溝がみられる。前正中裂と前外側溝との間の隆まりは(延髄)錐体ととよばれ、錐体路に相当する。錐体交叉は前正中裂を横切って走る線維として外表面からも認められる。前外側溝と後外側溝との間には背側と腹側に隆起がある。腹側の楕円形の隆起はオリーブで、中にはオリーブ核がある。背側の隆起は灰白結節で、三叉神経脊髄路と脊髄路核に相当する。後中間溝と後外側溝の間には楔状束核と薄束核に一致して、外側には楔状束結節と内側には薄束結節とがみられる。さらに上外方には下小脳脚が存在する。②横断面:脊髄との移行部の高さでは、中心管の背側には後索核(楔状束核と薄束核)がある。これらの核からの線維は内弓状線維となり腹内側に向かい交叉する(これを毛帯交叉とよぶ)。交叉後は錐体の背側に集まり内側毛帯を形成する。一方、腹側では錐体交叉をした線維が背外側の側索に入るのがみられる。オリーブ核の高さでは、一般に延髄背側部には基板および翼板由来の脳神経核が配列されている。内側から外側にかけて体性運動性の舌下神経核、一般内臓遠心性迷走神経背側核(内側核)と唾液核がある。同じく基板由来の特殊内臓遠心性の舌咽、迷走、副神経の疑核は腹外方に位置している。さらに、これらの外側には翼板由来の一般内臓感覚性の迷走神経背側核(外側核)、特殊内臓感覚性の孤束と孤束核があり、一般体性感覚性の三叉神経脊髄路核は延髄の最も外側に位置している。その他、副楔状束核が楔状束核の外側に、介在核が舌下神経核の外側にある。これらの神経核の腹側には網様体とよばれる。ここには縫線核がある。延髄背側部で縫線の両側には内側縦束が通り、その腹側には三角形の内側毛帯がある。腹側部には錐体とその背側から外側にオリーブ核がある。なお第四脳室底の中心灰白質の内側部を背側縦束が通る。延髄も上・下行性伝導路を通過させる。また、延髄には第8~12脳神経の諸核、呼吸中枢、循環中枢などが存在し、これらへの圧力(ヘルニア、頭蓋内圧亢進、などによるもの)は昏睡と死をまねく。延髄とはいうのは脊髄の延長部という意味で名づけられたらしい。このラテン名を最初に使ったのはハイステルLorenz Heister(1740)であるが、橋と脊髄の間の部位に限局して使いだしたのはハレルAlbrecht von Haller(1750)である。延髄はその膨らんだ感じから球とも呼ばれる。 延髄の発生 development of the medulla oblongata:菱脳の尾側半である菱脳は全体として延髄となる。 延髄の頭側約3分2は菱脳窩の尾側半を形成し、舌咽、迷走、副および舌下神経の諸核を生ずる。尾側約3分1の範囲では、内腔は第四脳室の形成に参加せず、狭い裂隙状の中心管として脊髄中心管につづく。この範囲では発生様式も内部構造も脊髄に類似するが、特別な者として翼板から後索核(薄束核と楔状束核)が生ずる。この核は脊髄後索に接続する中継核で、この核から出る神経線維は腹内方に走り、底板の縁帯で交叉した後、正中線の両側を上行する著明な線維束(内側毛帯)をつくり、視床に達する。 翼板と蓋板の移行部である菱脳唇から発生した細胞は腹内方に遊走して、基板の縁帯の中にはなはだ大きい神経核(オリーブ核)を形成する。この核から出る神経線維も基板の縁帯において交叉し、反対側の小脳に達する。このおうに底伴音縁帯は交叉線維で満たされて厚くなり正中縫線となる。 延髄においても上行および下行線維は、はじめは縁帯を通っている。しかし発生が進んで上行およびとくに下行線維が増えると、これらは外套層にも親友するようになる。また上述のように外套層の中には横走線維も多くなるので、はじめ比較的明瞭であった灰白質と白質の区別は次第に不明瞭となる。 胎生4ヶ月において延髄の腹側面で正中線の両側に接する部位の縁帯は、大脳皮質からの下行線維(錐体路線維)によって埋められ、これを延髄錐体と言うが、これは大脳皮質に属するもので、延髄固有の構造物ではない。)
- 831_10【Spinal cord脊髄 Medulla spinalis】 It extends from the end of the medulla oblongata near the exit of the first spinal nerve to the beginning of the terminal filum at L1 or L2.
→(脊髄は頚部(頚髄)、胸部(胸髄)、腰部(腰髄)、仙骨部または脊髄円錐(仙髄と尾髄)とからなり、それぞれ髄節に分かれ、それに対応して31対の脊髄神経が出る。頚髄では8対の頚神経、胸髄では12対の胸神経、腰仙髄では各々5対の腰神経と仙骨神経とが出る。尾髄からは通常1対の尾骨神経が出る。上肢および下肢支配の神経の出る頚髄下部と腰髄下部は発達が著しく、太くなっており、それぞれ頸膨大、腰部大とよばれる。脊髄下端は細くなり脊髄円錐となっておわる。その高さは成人では第1ないし第2腰椎の高さに相当する。新生児、幼児では低く第3腰椎の高さでおわっている。脊髄円錐の先はさらに細く糸状の終糸となって尾骨の背面に付着している。終糸に沿って走る脊髄神経の束はその形状から馬尾とよばれている。脊髄外側面でその腹側と背側の正中には(前)正中裂および(後)正中溝とよばれる溝があり、脊髄を左右の半分に分けている。前者は後者より深く、そこには前脊髄動脈が走っている。左右の脊髄半の外側面には腹側の前外側溝と背側の後外側溝の二つの溝がある。頚髄の高さの背側面は中心部の灰白質とその周辺の白質から成る。灰白質はそれぞれ前角(柱)、中間質(帯)、後角(柱)がある。灰白質の中央を貫いて中心管が通る。上方は第四脳室に開き、下方は脊髄炎水の所では拡大して終室となる。白質は前外側溝と後外側行と②より腹側の前索と外側の側索および背側の後索の3部分に分けられる。頚髄の高さで後索は後中間溝により内・外の薄束と楔状束とに分けられる。)
- 831_11Monro, Foramen of【Interventricular foramen室間孔 Foramen interventriculare】 Opening between the lateral and third ventricles behind the genu of the fornix.
→(モンロー孔ともよばれる。左右の側脳室と第三脳室とを結ぶ連絡孔。スコットランドの医学者Alexander Monro II (1733-1817)により、1797年に発見された。彼の名前はモンロー・リヒター線Monro-Richter lineにも残っている。)
- 831_12【Corpus striatum線条体[広義の] Corpus striatum】 Structure that is currently viewed as consisting of the putamen. caudate nucleus, pallidum, and fasciculi.
→(線条体は尾状核とレンズ核を意味する。レンズ核はさらに被殻と淡蒼球に区別される。このうち尾状核と被殻は終脳胞の腹外側に出現する神経節丘より同一の細胞群として発生し、その後、のちに発達してくる内包によって二つの部分に隔てられたものである。尾状核と被殻とは内包に横切って走る栓状の灰白質によって互いに連なり、特に前下方では両者は構造的にも同じ細胞構築をもっている。線条体という名称は内包を横切って尾状核と被殻を結んでいる灰白質によってできる縞目と、さらに、尾状核や被殻の中を走る有髄線維の小束によってできる縞目とに基づくものである。したがって、尾状核と被殻とをまとめて線条体Striatumとよび、淡蒼球をPallidumとよんで対比することが多い。解剖学用では、慣用されてきたStriatumという語とは異なる意味内容をもつ語として、Corpus striatumが採用されているが、日本名ではどちらも「線条体」である点は注意を要する。一方、淡蒼球の発生や細胞弧対句はStriatumとは異なる。淡蒼球は有髄神経線維に富むため黄灰白色を呈し、赤みを帯びた暗灰色のStriatumとは肉眼的にも明らかに識別できる。系統発生的視点に立って、尾状核と被殻を新線条体(Neostriatum)、淡蒼球を古線条体(Paleostriatum)、扁桃体を原線条体(Archistriatum)とよぶことがある。)
- 831_13【Anterior commissure前交連 Commissura anterior】 Commissure located anterior to the fornical column. It is readily visible in the anterior wall of the third ventricle.
→(間脳の前交連は第三脳室の前壁をつくる終板の後ろにある横走線維束である。前部は小線維束で、左右の両側の嗅脳系を結び、後部は大きな線維束で、左右両側の側頭葉に連絡する。前交連は小さな密な線維束で、脳弓柱の吻側で正中線を横切る。これは全体として自転車のハンドルに似た形態をしていて、肉眼標本では明瞭ではないが、2つの部分から構成される。前交連の小さい前部は肉眼標本ではあきらかでないが両側の嗅球を連絡している。大きい後部は主として両側の中側頭回および下側頭回の間を連絡する。)
- 831_14【Rhinencephalon; Olfactory brain嗅脳 Rhinencephalon】
→(嗅脳は嗅覚に直接関連する脳半球の部分を示す主に古典的な集合語、嗅球、嗅索、嗅三角、梁下野、前有孔質、終板傍回などからなる。嗅球、嗅索、嗅三角をまとめて嗅葉と呼ぶこともある。)
- 831_15【Optic stalk眼胞茎;眼茎;視茎 Pedunculus opticus】
→()
- 831_16【Optic chiasm; Optic chiasma視神経交叉;視交叉;視束交叉 Chiasma opticum; Chiasma fasciculorum opiticorum】 Decussation of medial optic nerve fibers between the optic tract and optic nerve.
→(視神経交叉は視床下部の漏斗の吻側にある扁平な線維板で、視神経線維が交叉しているところ。視交叉の背側から両側に開いて出る線維束は視索である。第三脳室の終板と灰白隆起の間で視交叉は第三脳室の底の一部を成す(視交叉陥凹)。視交叉はその上面で(終板の前方)前交連動脈と接し、下面はトルコ鞍の鞍隔膜の上に乗っている。眼球網膜の鼻側半からの線維は交叉して対側へ行き、側頭半からの線維は同側を交叉せずに後方へすすむ。下垂体前葉から発生する腫瘍が視交叉を圧迫することがある。)
- 831_17【Infundibulum of pituitary gland漏斗(下垂体の) Infundibulum】 Pituitary stalk.
→(下垂体の漏斗は視床下部の腹側方の突出部とその中にある第三脳室の陥凹によって形成される。漏斗の最も遠位に突出した部分が下垂体後葉(神経下垂体)であり、漏斗の突出部と正中隆起を結合する組織は漏斗柄とよばれる。)
- 831_18【Hypothalamic sulcus視床下溝 Sulcus hypothalamicus】 Furrow extending from the interventricular foramen to the entrance to the cerebral aqueduct. It divides the dorsal and ventral portions of the thalamus.
→(視床下溝は第三脳室両側の外側壁にある溝で視床内側面と視床下部の間にある境界溝に一致する。室間孔から後交連までいたる。)
- 831_19【Pineal gland; Pineal body松果体;松果腺 Glandula pinealis; Epiphysis cerebri; Corpus pineale】 It develops from the roof of the diencephalon and overlies the tectal plate.
→(松果体は円錐形の小体で、後交連の領域で、第三脳室の天井に付着している。これは痕跡的な腺であるらしく、血管の豊富な結合組織の小柱の網工からなり、その網眼の中には神経膠細胞および松果体細胞pinealocytesあるいはepiphysial cellがみられる。サルの松果体細胞はセロトニン(5-HT)とコレシストキニン(CCK)を含んでいる。哺乳類の松果体細胞は、系統発生学的には、感覚神経性光受容体の要素と関係しており、これは、分泌細胞になる物が多いが、間接的に光受容性をもっている。これらの突起の棍棒状をした終末は、血管血管を取り囲む血管周囲腔に接して終わる。松果体の分泌のうちで最もよく知られた物は、セロトニン、ノルアドレナリンおよびメラトニンという生体産生アミンであるが、その他に、サイロトロピン遊離ホルモン(TRH)、黄体形成ホルモン遊離ホルモン(LHRH)およびソマトスタチン(SRIF)のような、視床下部で形成されると同定されたペプチドを、かなりの濃度で含んでいる。セロトニンは、松果体細胞の中で合成されて細胞外の隙間に放出される。ノルアドレナリンは、松果体の実質細胞に終止ししている交感神経ニューロンの中で合成される。松果体は、昼間光の変動に敏感なN-アセチルトランスフェラーゼおよびヒドロキシインドール-o-メチルトランスフェラーゼという2つの酵素の働きによって、セロトニンからメラトニンを合成する。メラトニン合成の毎日の変動は周期性であり、光入力の毎日の周期に直接関係している。光は、日周期を環境の光周期に一致させる。そして、さらに、まだ確認されていない経路を経て、神経性信号が松果体に運ばれることを速やかに遮断するように働く。N-アセチルトランスフェラーゼの活性は、昼夜に高められるが、光にさらすと、酵素の活性が失われる。視床下部の視交叉上核を両側性に傷害すると、この核は網膜視床下部経路を受けているので、松果体のN-アセチルトランスフェラーゼにおける周期がなくなり、その結果、ヒドロキシインドール-o-メチルトランスフェラーゼの活性のレベルが下がる。そのような傷害によって、自発運動の活性の日周期および栄養補給と水飲み行動の両方の日周期がなくなる。雄のネズミでは、視交叉上核の傷害によって、正常な発情周期がなくなる。網膜視床下部路は、視交叉上核の構造との直接の相互作用によって、松果体のはたらきを変える。環境の光は、日周期をその周期に一致させる働きと伝達する働きをもっているとみなされ得る。内在性振動発生装置を光周期にのせるための光後下はゆっくりであるが、信号伝達に及ぼす光の後下は速やかである。そして、それによって、光によってN-アセチルトランスフェラーゼが速やかに“消失”することと、持続光によって日周期が妨げられることを、おそらくは説明できるであろう。視交叉上核からの単一神経路が、松果体によるメラトニン形成に関係する両酵素を調節しているが、これらの結合についての詳細は、なお、あきらかになっていない。間接的に証明された根拠は視交叉上核から松果体への神経路には、視床下部の灰白隆起の領域、内側前脳束および中間質外側路への中継路が含まれることを示唆している。このように、松果体は、交感神経ニューロンを経て受けた信号をメラトニンという内分泌物に変換させる、神経内分泌変換体ともいうべきものであるらしい。松果体の分泌は視床下部の働きを変化さえるが、それは、内分泌が心身の血液循環あるいは、脳脊髄液に入ってからののちに作用する。松果体のセロトニンとメラトニンの日内変動は、光入力の周期に応じて起こる。松果体活動のこれらの周期性変化は、この腺が生物学的時計としてはたらいて、生理学的過程と行動学的過程とを調節する信号を出していることを示唆している。これらの変動は日周期circadian rhythmsと呼ばれ、環境からの刺激が存在すれば、ほぼ24時間周期を示すだけである。松果体実質の腫瘍は、性的機能を低下させて思春期を送らせるが、他方、松果体を破壊するとしばしば早発思春期を伴う。これらの観察は、松果体が、性腺および生殖器系に抑制的影響を及ぼすことを示す実験的研究の結果と一致する。)
- 831_20【Tectal plate; Quadrigeminal plate蓋板;四丘体板;四丘板 Lamina tecti; Lamina quadrigemina】
→()
- 831_21【Cerebellum小脳 Cerebellum】 Part of the brain situated above the rhomboid fossa.
→(Cerebellumは、「大脳、脳」を意味するcerebrumの指小形で、「小さい脳」という意味である。Cerebrumは、「頭」を意味するギリシャ語のkararan由来する。 小脳は筋、関節などの深部組織、前庭、視覚、聴覚系などからの入力を直接あるいは間接的に受け、眼球運動を含む身体の運動調節を司る。小脳は正中部の虫部と外側部の小脳半球とに分けられる。いずれも多数の小脳溝により小脳回に細分される。この中、特定の小脳溝は深く、これにより小脳回の集合ができる。これを小脳小葉とよぶ。ヒトでは小脳は深い水平裂により上面と下面とに分けられ、虫部とそれに対応する半球に九つの小葉が区別される。系統発生的には小脳は前葉、後葉、片葉小節葉の3部分に分けられる。前葉は系統発生的に古く古小脳(Paleocerebellum)ともよばれ、脊髄小脳路、副楔状束核小脳路、オリーブ小脳路の一部、網様体小脳路などをうける。後葉は系統発生的に新しく、新小脳(Neocerebellum)とよばれる。とくに半球部は虫部より新しく、橋核、主オリーブ核などを介して大脳皮質と結合している。前葉と後葉とは第1裂により境される。片葉小節葉は原小脳(Archicerebellum)とよばれ最も古く前庭系との結合が著明である。後葉とは後外側裂で境される。後葉には虫部錐体と虫部垂との間に第2裂がある。ヒトの小脳小葉の形は他の動物のものと大きく異なりる。小脳全体は灰白質と白質とからなる。灰白質には小脳皮質と小脳核とがある。小脳皮質は小脳小葉の表面をなし遠心性軸索を出すPurkinje細胞と皮質内での結合を行う細胞とからなる。小脳核は深部にあり、室頂核、球状核、栓状核、歯状核の4核からなる。小脳皮質にはその結合から三つの縦帯が認められる。すなわち、正中部の虫部皮質、外側部の半球皮質および両者の境界部の虫部傍皮質である。虫部皮質のPurkinje細胞は室頂核に、虫部傍皮質は球状核と栓状核に、半球皮質は歯状核に投射する。小脳の中心部の白質塊は髄体とよばれ、遠心性および求心性繊維から出来ている。ここからは白質が分枝して(白質板)、小葉に分かれる。全体として樹の枝のようにみえるので、小脳活樹と名づけられている。小脳は三つの小脳脚により、延髄、橋、中脳と結合している。これは小脳の遠心路および求心路の通路となっている。 小脳の発生 development of the cerebellum:小脳は後脳の菱脳唇から発生する。後脳の菱脳唇は翼板の背外側につづく背内方に突出する高まりで、胎生2ヶ月の後半において急速に増大し、小脳板とよばれるようになる。左右の小脳板の間には菱脳蓋の頭側半分が介在するので、頭側部では左右の小脳板は相接しているが、尾側部では広く離れている。菱脳の中央部を頂点とする橋弯曲が高度になると、この部の菱脳蓋の左右方向の拡大によって、左右の小脳板の尾側部はいよいよ高度に引き離され、左右の小脳板は菱脳の長軸に直角な一直線をなすようになる。これと同時に左右の小脳板の頭側部(今では内側部)が合一するので、結局、正中部が小さくて左右両部が大きい単一の小脳原基が成立する。正中部からは小脳虫部が、左右両部からは小脳半球が形成される。 増大していく小脳原基の背側部には、やがて中部から半球に向かって走る溝が次々に出現して小脳を区画する。胎生5ヶ月のおわりには小脳虫部における10個の主な区分(小脳葉)がほぼ完成する。これらの小脳葉はそれぞれ固有の発育を行うが、その間に第2次、第3次の溝が生じて、各小脳葉を多数の小脳回に分ける。このような形態発生の結果広大な表面積を獲得した小脳の表面には小脳皮質とよばれる特別な灰白質が形成され、これに出入りする神経線維はその深部に集まって小脳白質を形成する。 小脳原基においても菱脳室に接する内側から表面に向かって胚芽層・外套層・縁帯の3層が分化する。胚芽層は神経が細胞をつくりだすが、胚芽層から発生するのは小脳核の神経細胞と小脳皮質のPurkinje細胞およびGolgi細胞である。小脳原基が3層に分化するとまもなく、外套層の表層部にやや大型の神経芽細胞が出現し、小脳板の背側面(表面)に平行に1列にならぶ。これがPurkinje細胞の幼若型である。ついで小脳板の尾側端部の胚芽層でさかんな細胞分裂がおこり、ここで生じた未分化細胞は縁帯の表層部を頭方に遊走して、小脳原位の全表面をおおう未分化細胞層を形成する。これを胎生顆粒層という。 胎生顆粒層の細胞は胚芽層における細胞分裂が終わるころから活発な分裂を開始し、神経細胞をつくりだす。この神経細胞は縁帯およびPurkinje細胞の層を貫いて、Purkinje細胞の層の下に達し、ここに新しい細胞層(内顆粒層)をつくる。胎生顆粒層からは、このほかに縁帯の中に散在する籠細胞や小皮質細胞が生ずる。必要な数の神経細胞を送り出すと胎生顆粒層における分裂はやみ、本層は速やかに消失する。一方、Purkinje細胞は縁帯の中に多数の樹状突起を伸長させる。縁帯はPurkinje細胞の樹状突起で満たされて厚くなり、核をあまり多く含まな灰白層となる。このようにして小脳の全表面は、表面から灰白層・Purkinje細胞層・内顆粒層の3層から成る小脳皮質でおおわれることになる。)