Spalteholz HANDATLAS DER ANATOMIE DES MENSCHEN VON WERNER SPALTEHOLZ
メニューは解剖学(TA)にリンクしてあります。図の番号をクリックすると下記の説明へ、右側の用語をクリックすると解剖学(TA)にジャンプします。
855


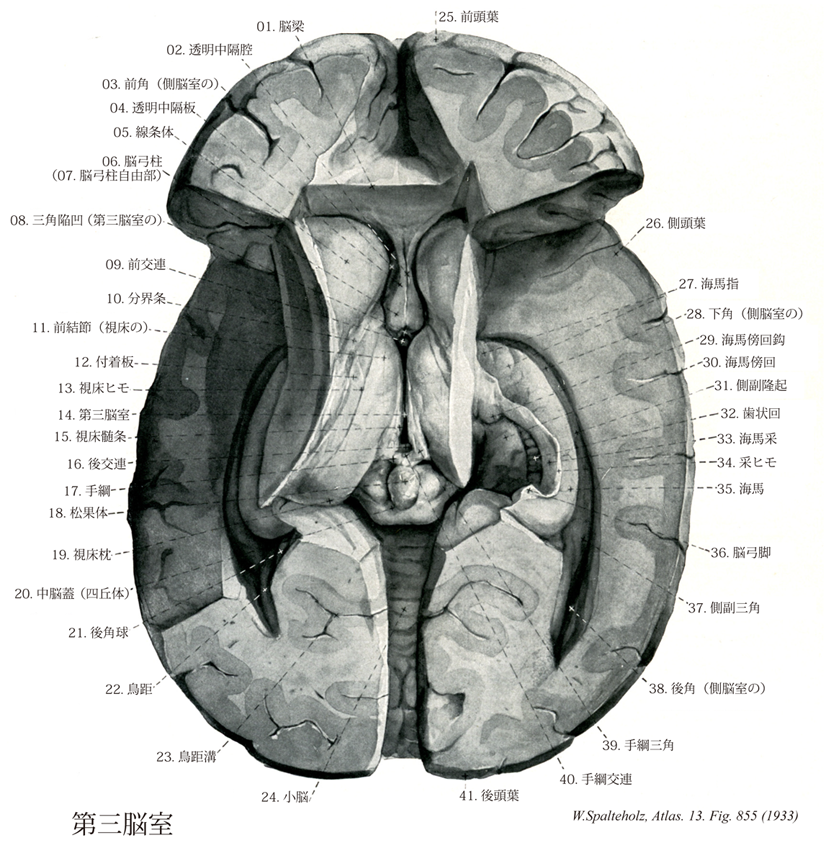
- 855_01【Corpus callosum脳梁 Corpus callosum】 Transverse nerve fibers connecting the two cerebral hemispheres at the base of the longitudinal cerebral fissure.
→(脳梁は左右の大脳皮質、ことに新皮質を結合する線維の集合したもので、系統発生的には最も新しく、ヒトでは非常に発育がよい。その前後経はほぼ7.7cmである。脳梁は正中断では全体としては釣針状で、4つの部分が区別される。後端部は膨大し、脳梁膨大といい、その前方に続いて水平に走る部分を脳梁幹とよぶ脳梁はその前端では強く屈曲し、脳梁膝をつくる。これはさらに後下方にくちばしのように尖って脳梁吻となり、しだいに薄くなって終板に続く。)
- 855_02【Cave of septum pellucidum透明中隔腔 Cavum septi pellucidi】 Enclosed cavity of variable size between the two laminae.
→(透明中隔腔は左右の透明中隔板の間に挟まれた大きさ不定の腔。ヒトでは10%以下の率で存在し、第3脳室に連なる。)
- 855_03【Frontal horn of lateral ventricle; Anterior horn of lateral ventricle前角;前頭角(側脳室の) Cornu frontale ventriculi lateralis; Cornu anterius ventriculi lateralis】 Portion extending anteriorly from the interventricular foramen. It is bounded medially by the septum pellucidum, laterally by the head of caudate nucleus, superiorly by the body of corpus callosum, and anteriorly and inferiorly by the genu and rostrum of corpus callosum, respectively.
→(側脳室の前角は室間孔より前方の部分、内側壁は透明中隔、外側壁は尾状核頭、前壁及び上下壁は脳梁によってつくられる。)
- 855_04【Lamina of septum pellucidum透明中隔板 Lamina septi pellucidi】 Paired layer forming the septum pellucidum. It forms the lateral wall of the cave of septum pellucidum.
→(透明中隔板は透明中隔の一対の板。透明中隔腔の側壁をなす。)
- 855_05【Corpus striatum線条体[広義の] Corpus striatum】 Structure that is currently viewed as consisting of the putamen. caudate nucleus, pallidum, and fasciculi.
→(線条体は尾状核とレンズ核を意味する。レンズ核はさらに被殻と淡蒼球に区別される。このうち尾状核と被殻は終脳胞の腹外側に出現する神経節丘より同一の細胞群として発生し、その後、のちに発達してくる内包によって二つの部分に隔てられたものである。尾状核と被殻とは内包に横切って走る栓状の灰白質によって互いに連なり、特に前下方では両者は構造的にも同じ細胞構築をもっている。線条体という名称は内包を横切って尾状核と被殻を結んでいる灰白質によってできる縞目と、さらに、尾状核や被殻の中を走る有髄線維の小束によってできる縞目とに基づくものである。したがって、尾状核と被殻とをまとめて線条体Striatumとよび、淡蒼球をPallidumとよんで対比することが多い。解剖学用では、慣用されてきたStriatumという語とは異なる意味内容をもつ語として、Corpus striatumが採用されているが、日本名ではどちらも「線条体」である点は注意を要する。一方、淡蒼球の発生や細胞弧対句はStriatumとは異なる。淡蒼球は有髄神経線維に富むため黄灰白色を呈し、赤みを帯びた暗灰色のStriatumとは肉眼的にも明らかに識別できる。系統発生的視点に立って、尾状核と被殻を新線条体(Neostriatum)、淡蒼球を古線条体(Paleostriatum)、扁桃体を原線条体(Archistriatum)とよぶことがある。)
- 855_06【Column of fornix脳弓柱 Columna fornicis】
→(脳弓柱は没部と出部からでる。没部は左右の乳頭体から始まり、視床下部内を前上方に走り、前交連の後ろで出部に移行する。出部は大脳半球の正中断面で見える部分で、上行するとともにしだいに左右が互いに近づき、ついで後背側方に走り、前方は透明中隔板と癒着している。出部は脳梁幹の下で脳弓体に移行する。)
- 855_07【Free part of column of fornix脳弓柱自由部;脳弓柱出部 Pars libera columnae fornicis】
→()
- 855_08【Triangular recess of third ventricle三角陥凹(第三脳室の) Recessus triangularis (Ventriculus tertius)】
→()
- 855_09【Anterior commissure前交連 Commissura anterior】 Commissure located anterior to the fornical column. It is readily visible in the anterior wall of the third ventricle.
→(間脳の前交連は第三脳室の前壁をつくる終板の後ろにある横走線維束である。前部は小線維束で、左右の両側の嗅脳系を結び、後部は大きな線維束で、左右両側の側頭葉に連絡する。前交連は小さな密な線維束で、脳弓柱の吻側で正中線を横切る。これは全体として自転車のハンドルに似た形態をしていて、肉眼標本では明瞭ではないが、2つの部分から構成される。前交連の小さい前部は肉眼標本ではあきらかでないが両側の嗅球を連絡している。大きい後部は主として両側の中側頭回および下側頭回の間を連絡する。)
- 855_10【Stria terminalis分界条 Stria terminalis】 Longitudinal band of efferent fibers from the amygdaloid body. It is accompanied by the superior thalamostriate vein in the angle between the thalamus and caudate nucleus.
→(分界条は背側視床と尾状核の間にある狭い白質で、主として扁桃体から起こり、脳弓と平行して走る。この線維の中には視索前野と視床下部と腹内側核でシナプス結合するものがある。)
- 855_11【Anterior thalamic tubercle前結節(視床の);視床前結節 Tuberculum anterius thalami】 Small protuberance on the anterior end of the thalamus. Attachment site of the stria medullaris of the thalamus.
→(視床の前結節は視床前端の小結節。背側視床の前面は狭く、その背側部は前方にやや突出し、視床前結節と呼ばれる。)
- 855_12【Lamina affixa付着板 Lamina affixa】 Floor of the lateral ventricle between the stria terminalis and choroid line.
→(付着板は発生後期には視床上面に付着して側脳室の中心部の床を形成する。側脳室の内面をおおう上衣層の一部にすぎないが、二次的に間脳に属する視床の背側面の外側部を多い、これと癒着したものである。)
- 855_13【Taenia thalami視床ヒモ;視床脈絡ヒモ Taenia thalami; Taenia chorioidea thalami】 Upper margin of the stria medullaris of thalamus. Attachment site of the choroid plexus of third ventricle.
→(第三脳室の薄い上皮性の天井は視床の比較的鋭い上縁に付着している(視床ヒモ)。この視床ヒモに沿って視床髄条(中隔部その他と手綱核を連絡する神経線維束)が走る。視床ヒモは背側方では手綱三角に達しており、そこで手綱交連に沿って正中線を越え、第三脳室の上皮性の天井を閉じている。視床ヒモは背側方ではMonro孔を通過して脈絡ヒモに連続している。)
- 855_14【Third ventricle第三脳室 Ventriculus tertius】 Diencephalic part of the cerebral ventricular system. It extends from the lamina terminalis to the cerebral aqueduct.
→(第三脳室は左右の間脳の間にある背腹方向にスリット状を示す腔である。前壁は終板と前交連によってつくられる。前上部には室間孔が開口し、左右の側脳室と交通し、後方は中脳水道と連絡する。後壁は松果体に入り込む松果体陥凹がみられ、下壁は視床下部によってつくられ、視交叉陥凹、漏斗陥凹がみられる。外側壁を形成している視床と視床下部の境には視床下溝が走る。なお、脳室の前上方部に第三脳室終脳部とよばれる部分がある。)
- 855_15【Stria medullaris of thalamus視床髄条 Stria medullaris thalamica】 Bundle of white fibers running from the anterior thalamic tubercle to the habenula between the dorsal and medial surfaces of the thalamus.
→(視床髄条は視床ヒモの下にある細長く稠密な線維束。視床髄条は後方では手綱に移行し、前有孔質や嗅三角などから起こって内側および外側手綱核に終わる線維から成る(嗅手綱路)。)
- 855_16【Posterior commissure後交連 Commissura posterior; Commissura epithalamica】 Commissure situated between the pineal recess and the opening of cerebral aqueduct. Crossing site of fibers from the surrounding area.
→(後交連は、中脳と間脳の背側における境界をなす。この小さな交連は中脳水道と第三脳室の移行部で上丘の上方の中心灰白質の背側にある。交連線維は外側で扇状にひろがり、その周囲を後交連核と総称される細胞が取り囲む。これまで明らかにされた後交連の構成線維は、①視蓋前核、②後交連核、③間質核からの線維がある。瞳孔の対光反射に関与する線維は後交連で交叉する。後交連の下方の中脳水道上衣は線毛をもつ高い柱状細胞よりなる。このような変形上衣細胞は交連下器官を形成し、分泌機能をもち、脳室周囲器官の一つとされている。交連下器官は中脳において脳血液関門をもたない唯一の部分である。)
- 855_17【Habenula手綱 Habenula】 Synaptic site between the pineal gland, brainstem, and olfactory centers.
→(手綱は第三脳室後壁の背側面において、松果体の吻側を横走する索状の白質を手綱と呼ぶ(松果体の「手綱」とう意味)。手綱の神経線維は正中部で交叉して手綱交連を形成する。手綱の外側端は三角状に広がっており手綱三角と呼ばれ、その吻側より視床髄条が入る。手綱三角の深部には手綱核が存在する。手綱核には、小形神経細胞の密集する内側手綱核と、比較的大きい神経細胞が比較的疎に集まっている外側手綱核が区別される。手綱核からは反屈束がおこり腹側の脚間核に向かう。反屈束を構成する神経線維のうち、中心部を走る細い線維は主として内側手綱核からおこり脚間核に終止する(手綱脚間線維)。一方、反屈束の周辺部を構成する比較的太い神経線維は主として外側手綱核からおこり、脚間核の背外側部を通過して、中脳の背側被蓋核と腹側被蓋核に終止する。また、反屈束には以上のような神経線維のほか、前有孔質や視床下部などの前脳の底部領域よりおこり、視床髄条を通って手綱核にいたり、さらに手綱核を通過して反屈束に加わる線維や、反屈束を上行する線維なども含まれている。)
- 855_18【Pineal gland; Pineal body松果体;松果腺 Glandula pinealis; Epiphysis cerebri; Corpus pineale】 It develops from the roof of the diencephalon and overlies the tectal plate.
→(松果体は円錐形の小体で、後交連の領域で、第三脳室の天井に付着している。これは痕跡的な腺であるらしく、血管の豊富な結合組織の小柱の網工からなり、その網眼の中には神経膠細胞および松果体細胞pinealocytesあるいはepiphysial cellがみられる。サルの松果体細胞はセロトニン(5-HT)とコレシストキニン(CCK)を含んでいる。哺乳類の松果体細胞は、系統発生学的には、感覚神経性光受容体の要素と関係しており、これは、分泌細胞になる物が多いが、間接的に光受容性をもっている。これらの突起の棍棒状をした終末は、血管血管を取り囲む血管周囲腔に接して終わる。松果体の分泌のうちで最もよく知られた物は、セロトニン、ノルアドレナリンおよびメラトニンという生体産生アミンであるが、その他に、サイロトロピン遊離ホルモン(TRH)、黄体形成ホルモン遊離ホルモン(LHRH)およびソマトスタチン(SRIF)のような、視床下部で形成されると同定されたペプチドを、かなりの濃度で含んでいる。セロトニンは、松果体細胞の中で合成されて細胞外の隙間に放出される。ノルアドレナリンは、松果体の実質細胞に終止ししている交感神経ニューロンの中で合成される。松果体は、昼間光の変動に敏感なN-アセチルトランスフェラーゼおよびヒドロキシインドール-o-メチルトランスフェラーゼという2つの酵素の働きによって、セロトニンからメラトニンを合成する。メラトニン合成の毎日の変動は周期性であり、光入力の毎日の周期に直接関係している。光は、日周期を環境の光周期に一致させる。そして、さらに、まだ確認されていない経路を経て、神経性信号が松果体に運ばれることを速やかに遮断するように働く。N-アセチルトランスフェラーゼの活性は、昼夜に高められるが、光にさらすと、酵素の活性が失われる。視床下部の視交叉上核を両側性に傷害すると、この核は網膜視床下部経路を受けているので、松果体のN-アセチルトランスフェラーゼにおける周期がなくなり、その結果、ヒドロキシインドール-o-メチルトランスフェラーゼの活性のレベルが下がる。そのような傷害によって、自発運動の活性の日周期および栄養補給と水飲み行動の両方の日周期がなくなる。雄のネズミでは、視交叉上核の傷害によって、正常な発情周期がなくなる。網膜視床下部路は、視交叉上核の構造との直接の相互作用によって、松果体のはたらきを変える。環境の光は、日周期をその周期に一致させる働きと伝達する働きをもっているとみなされ得る。内在性振動発生装置を光周期にのせるための光後下はゆっくりであるが、信号伝達に及ぼす光の後下は速やかである。そして、それによって、光によってN-アセチルトランスフェラーゼが速やかに“消失”することと、持続光によって日周期が妨げられることを、おそらくは説明できるであろう。視交叉上核からの単一神経路が、松果体によるメラトニン形成に関係する両酵素を調節しているが、これらの結合についての詳細は、なお、あきらかになっていない。間接的に証明された根拠は視交叉上核から松果体への神経路には、視床下部の灰白隆起の領域、内側前脳束および中間質外側路への中継路が含まれることを示唆している。このように、松果体は、交感神経ニューロンを経て受けた信号をメラトニンという内分泌物に変換させる、神経内分泌変換体ともいうべきものであるらしい。松果体の分泌は視床下部の働きを変化さえるが、それは、内分泌が心身の血液循環あるいは、脳脊髄液に入ってからののちに作用する。松果体のセロトニンとメラトニンの日内変動は、光入力の周期に応じて起こる。松果体活動のこれらの周期性変化は、この腺が生物学的時計としてはたらいて、生理学的過程と行動学的過程とを調節する信号を出していることを示唆している。これらの変動は日周期circadian rhythmsと呼ばれ、環境からの刺激が存在すれば、ほぼ24時間周期を示すだけである。松果体実質の腫瘍は、性的機能を低下させて思春期を送らせるが、他方、松果体を破壊するとしばしば早発思春期を伴う。これらの観察は、松果体が、性腺および生殖器系に抑制的影響を及ぼすことを示す実験的研究の結果と一致する。)
- 855_19【Pulvinar of thalamus視床枕 Pulvinar thalami】 Posterior, freely projecting portion of the thalamus.
→(視床枕は視床の後部と背外側を形成する大きな灰白質塊で、これの尾方は、内側膝状体、外側膝状体および中脳の背外側面の上に張り出している。視床枕は視床枕核群ともよばれており細胞学的にはかなり均一であるので、局所的な位置関係を基にして細分される。視床枕を形成するのは、明るく染まった、中等度の大きさの、多極性の細胞で、それらの細胞の分布密度と配列は、視床枕の部位によって異なっている。視床枕の前部の細胞は小さく、明るく染まり、散在性に配列している。下部は、視床枕の主部から、上丘腕の神経線維によって隔てられており、散在性の濃染する細胞で構成される。外側部を、外側髄板から広がる斜めの線維束が横切っている。視床枕の諸核は、長い、上行性の感覚神経路からは入力を受けてはいないが、その下部は、上丘の浅層のいくつかの層からの投射を受けている。局所的には、この投射は、反対側の視野の半分に相当する。視床枕の下部とそれい隣接する外側部とは、線条野を含む後頭葉の皮質と相互に結合する。視床枕の下部とそれに隣接する外側部はそれぞれ、反対側の視覚野の半分が復元され、網膜の部位局在的に、次の各部に投射している。すなわち、①皮質の18野と19野と、②有線野(17野)で、そこでは、線維が、顆粒層の上にある諸層に終止する。これらの結果によって、3つの視覚局在性をもった系統の入力が、視床(外側膝状体、視床枕の下部、およびそれに隣接する視床枕の外側部)から、一次視覚野に達し、しかも、そのいろいろな層に終止することが明らかになった。視床枕の下部から17野、18野および19野への投射は、膝状体外視覚神経路の中の最後の連絡を形成する。(視床枕の下核に隣接している部分以外の)視床枕の外側核は、側頭葉に投射し、同じ領域と相互に連絡している。視床枕の内側部は、上側頭回に投射しているらしい。)
- 855_20【Tectum of midbrain中脳蓋;四丘体 Tectum mesencephali; Corpora quadrigemina】 Part of the mesencephaIon lying on the tegmentum of midbrain.
→(中脳蓋は中脳水道が通り、屋根状になった脳背側部。上丘と下丘が含まれる。)
- 855_21【Bulb of occipital horn of lateral ventricle後角球;後頭角球(側脳室の) Bulbus cornus posterioris ventriculi lateralis】 Eminence on the medial side of the posterior horn produced by fibers from the splenium of corpus callosum.
→(後角球は側脳室の後角内側面の背部にある丸い高まりで、大鉗子によってつくられた隆起。脳梁膨大中にある線維による。)
- 855_22【Calcarine spur鳥距 Calcar avis】 Elevation on the medial side of the posterior horn produced by the calcarine sulcus.
→(鳥距は後角内側面の2個の隆起の下方のもので、鳥距溝の沈下によって生じたもの。)
- 855_23【Calcarine sulcus; Calcarine fissure鳥距溝;鳥距裂 Sulcus calcarinus; Fissura calcarina】 Deep groove beneath the cuneus in the region of primary visual perception.
→(鳥距溝は大脳半球の内側面で、後頭極の近くから前方に向かってほぼ水平に走る深い溝である。前端で頭頂後頭溝に合する。一次視覚野は視覚中枢で、後頭葉の内側面において、鳥距溝の両側(Brodmannの17野)にある。視床の外側膝状体からの線維が視放線をつくってここに達する。視覚野の皮質は他の皮質部に比べて薄く(約1.5mm)、皮質内に表面に平行に走る有髄線維束(ジェンナリ線Gennari's strai or line)が発達し、これが肉眼的に認められるので有線野area striataといわれる。一側の視覚野には、同側の網膜耳側半部と反対側の網膜尾側半部とからの入力が復元される。また、網膜の上半分は鳥距溝の上方に、網膜の下半分は鳥距溝の下方に復元される。黄斑(網膜の後端で、視覚が最も鋭い部)は視覚野(17野)の後約3分の1部を占める広い領域に復元される。)
- 855_24【Cerebellum小脳 Cerebellum】 Part of the brain situated above the rhomboid fossa.
→(Cerebellumは、「大脳、脳」を意味するcerebrumの指小形で、「小さい脳」という意味である。Cerebrumは、「頭」を意味するギリシャ語のkararan由来する。 小脳は筋、関節などの深部組織、前庭、視覚、聴覚系などからの入力を直接あるいは間接的に受け、眼球運動を含む身体の運動調節を司る。小脳は正中部の虫部と外側部の小脳半球とに分けられる。いずれも多数の小脳溝により小脳回に細分される。この中、特定の小脳溝は深く、これにより小脳回の集合ができる。これを小脳小葉とよぶ。ヒトでは小脳は深い水平裂により上面と下面とに分けられ、虫部とそれに対応する半球に九つの小葉が区別される。系統発生的には小脳は前葉、後葉、片葉小節葉の3部分に分けられる。前葉は系統発生的に古く古小脳(Paleocerebellum)ともよばれ、脊髄小脳路、副楔状束核小脳路、オリーブ小脳路の一部、網様体小脳路などをうける。後葉は系統発生的に新しく、新小脳(Neocerebellum)とよばれる。とくに半球部は虫部より新しく、橋核、主オリーブ核などを介して大脳皮質と結合している。前葉と後葉とは第1裂により境される。片葉小節葉は原小脳(Archicerebellum)とよばれ最も古く前庭系との結合が著明である。後葉とは後外側裂で境される。後葉には虫部錐体と虫部垂との間に第2裂がある。ヒトの小脳小葉の形は他の動物のものと大きく異なりる。小脳全体は灰白質と白質とからなる。灰白質には小脳皮質と小脳核とがある。小脳皮質は小脳小葉の表面をなし遠心性軸索を出すPurkinje細胞と皮質内での結合を行う細胞とからなる。小脳核は深部にあり、室頂核、球状核、栓状核、歯状核の4核からなる。小脳皮質にはその結合から三つの縦帯が認められる。すなわち、正中部の虫部皮質、外側部の半球皮質および両者の境界部の虫部傍皮質である。虫部皮質のPurkinje細胞は室頂核に、虫部傍皮質は球状核と栓状核に、半球皮質は歯状核に投射する。小脳の中心部の白質塊は髄体とよばれ、遠心性および求心性繊維から出来ている。ここからは白質が分枝して(白質板)、小葉に分かれる。全体として樹の枝のようにみえるので、小脳活樹と名づけられている。小脳は三つの小脳脚により、延髄、橋、中脳と結合している。これは小脳の遠心路および求心路の通路となっている。 小脳の発生 development of the cerebellum:小脳は後脳の菱脳唇から発生する。後脳の菱脳唇は翼板の背外側につづく背内方に突出する高まりで、胎生2ヶ月の後半において急速に増大し、小脳板とよばれるようになる。左右の小脳板の間には菱脳蓋の頭側半分が介在するので、頭側部では左右の小脳板は相接しているが、尾側部では広く離れている。菱脳の中央部を頂点とする橋弯曲が高度になると、この部の菱脳蓋の左右方向の拡大によって、左右の小脳板の尾側部はいよいよ高度に引き離され、左右の小脳板は菱脳の長軸に直角な一直線をなすようになる。これと同時に左右の小脳板の頭側部(今では内側部)が合一するので、結局、正中部が小さくて左右両部が大きい単一の小脳原基が成立する。正中部からは小脳虫部が、左右両部からは小脳半球が形成される。 増大していく小脳原基の背側部には、やがて中部から半球に向かって走る溝が次々に出現して小脳を区画する。胎生5ヶ月のおわりには小脳虫部における10個の主な区分(小脳葉)がほぼ完成する。これらの小脳葉はそれぞれ固有の発育を行うが、その間に第2次、第3次の溝が生じて、各小脳葉を多数の小脳回に分ける。このような形態発生の結果広大な表面積を獲得した小脳の表面には小脳皮質とよばれる特別な灰白質が形成され、これに出入りする神経線維はその深部に集まって小脳白質を形成する。 小脳原基においても菱脳室に接する内側から表面に向かって胚芽層・外套層・縁帯の3層が分化する。胚芽層は神経が細胞をつくりだすが、胚芽層から発生するのは小脳核の神経細胞と小脳皮質のPurkinje細胞およびGolgi細胞である。小脳原基が3層に分化するとまもなく、外套層の表層部にやや大型の神経芽細胞が出現し、小脳板の背側面(表面)に平行に1列にならぶ。これがPurkinje細胞の幼若型である。ついで小脳板の尾側端部の胚芽層でさかんな細胞分裂がおこり、ここで生じた未分化細胞は縁帯の表層部を頭方に遊走して、小脳原位の全表面をおおう未分化細胞層を形成する。これを胎生顆粒層という。 胎生顆粒層の細胞は胚芽層における細胞分裂が終わるころから活発な分裂を開始し、神経細胞をつくりだす。この神経細胞は縁帯およびPurkinje細胞の層を貫いて、Purkinje細胞の層の下に達し、ここに新しい細胞層(内顆粒層)をつくる。胎生顆粒層からは、このほかに縁帯の中に散在する籠細胞や小皮質細胞が生ずる。必要な数の神経細胞を送り出すと胎生顆粒層における分裂はやみ、本層は速やかに消失する。一方、Purkinje細胞は縁帯の中に多数の樹状突起を伸長させる。縁帯はPurkinje細胞の樹状突起で満たされて厚くなり、核をあまり多く含まな灰白層となる。このようにして小脳の全表面は、表面から灰白層・Purkinje細胞層・内顆粒層の3層から成る小脳皮質でおおわれることになる。)
- 855_25【Frontal lobe前頭葉 Lobus frontalis】 Portion extending from the frontal pole to the central sulcus.
→(前頭葉は中心溝の前、そして外側溝の上にある。前頭葉の上外側表面は3つの脳溝によって4つの脳回に分けられる。前頭葉には、1次運動野はBrodmannの脳区分でいうと領域4(中心前回から中心傍小葉)を中心に錐体外路系の中枢があり、身体の反対側の随意運動を起こす。Betzの巨大細胞が特徴的であるが、この細胞からの線維は皮質脊髄路の3%程である。運動前野(2次運動野)はBrodmannの領域6,8,44など(中心前回の前部から上・中・下前頭回後部)にある錐体外路系の運動中枢。この部は一次運動野の活動プログラム化に働き、障害されると習得した運動が障害される。通常の運動障害はなく、失行と呼ばれる。前頭眼野は中心前回で、顔面支配領域の前方に位置する(主に8野で6,9野の一部)。眼球や眼瞼の共同運動の中枢である。補足運動野は上前頭回内部に位置し、姿勢や運動開始と関係するらしい。運動性言語中枢はBrodmannの領域44,45(三角部)に位置する。Brocaの言語野ともいい、言語発声に必要な口から口頭の筋を統合支配する中枢で、運動野と連絡して発声運動を行うという。この部が障害されると意味のある言語を発声できなくなる運動失語が生じる。運動前野は大脳皮質の前頭葉の前方を広く占有している連合野である(Brodmannの9,10野)。前頭前野は脳の極めて広範囲から上方を集めて行動のプログラミングを行う。靴紐を結んだり、ボタンをかけるなどの学習・経験による複雑な組織化された運動の遂行と関係がある。背側運動前野は運動の企画や準備に対応し、腹側運動前野は物体を認知して動作へ変換する情報に変えるといわれる。)
- 855_26【Temporal lobe側頭葉 Lobus temporalis】 Lobe bounded superiorly by the lateral sulcus.
→(側頭葉は外側溝より下方にある部分で、上外側面から下面におよび、後方は後頭葉および頭頂葉に移行する。上外側面では溝としては前後に走る上および下側頭溝があり、これらによって上、中および下側頭回が区画される。上側頭回の背側面で外側溝にかくれた部分には3本の横側頭溝があり、これらにより区別される二つの横側頭回(Heschl)がある。ここに聴覚野がある。これらの大部分は側頭葉下面皮質とともに連合中枢(側頭連合野)と目される。優位半球(主に左脳)の上側頭回の後部から角回にかけて感覚性言語中枢(Wernicke野)があるとされる。側頭葉下面では溝として前後に走る後頭側頭溝およびその内側をほぼこれと平行に走る側副溝がある。また側副溝の前方の延長部には浅い嗅脳溝がある。これは海馬傍回の前部と側頭葉の残部とを境する。回としては外側後頭側頭回は大脳上外側面における下側頭回が下面へ直接移行したもので、これらの両回の間を境する溝はない。内側後頭側頭回は後頭側頭溝と側副溝の間にあり、舌状回は側副溝後部と鳥距溝の間にあり、むしろ大部分は後頭葉に属する。)
- 855_27【Hippocampal digitationes; Hippocampal digitations海馬指;海馬指状突起 Digitationes hippocampi】 Claw-like projections of the pes hippocampi.
→(海馬の腹側部は大きく指のような隆起があるので、腹側部を海馬足、指のような隆起を海馬指とよんでいる。)
- 855_28【Inferior horn of lateral ventricle; Temporal horn of lateral ventricle;下角;側頭角;側頭部(側脳室の) Cornu temporale ventriculi lateralis; Cornu inferius ventriculi lateralis】 It extends with the hippocampus laterally and contains the choroid plexus.
→(側脳室の下角(側頭角)は側頭葉に向かって前下方に突出した部分で、この上外側壁は主として脳梁膨大から放散する脳梁線維からなり、これを壁板という。下壁には側副溝によるたかまり、すなわち側副隆起があり、その後方は三角形をなし、側副三角と呼ばれ、後角まで延びている。内側壁は複雑で、上壁との境界部に尾状核尾および分界条があり、その下に上衣層によっておおわれた側脳室脈絡叢があり、さらにその下には脳弓脚の続きである海馬采がある。)
- 855_29【Uncus; Uncus of parahippocampal gyrus; Uncus of hippocampal gyrus鈎;海馬傍回鈎;海馬鈎 Uncus; Uncus gyri parahippocampi; Uncus gyri hippocampi】 Anterior hook-shaped end of the parahippocampal gyrus.
→(鈎は海馬傍回の前端で、側頭葉底内側面に鈎でかかったようになっているところ。その前面は嗅皮質に、腹側面は内嗅野(entorhinal area)にそれぞれ相当する。深部には扁桃体がある。)
- 855_30【Parahippocampal gyrus; Hippocampal gyrus海馬傍回;海馬回 Gyrus parahippocampalis; Gyrus hippocampi】 Convolution situated below the hippocampal sulcus.
→(海馬傍回は海馬溝の下方、側副溝との間に大きな回である。海馬傍回の皮質の細胞構築は海馬台前部から順次海馬台、前海馬台を経て海馬体、歯状回にいたる間に次第に6層から3層構造に移り変わる。嗅内領(28野)は6層構造の皮質であるが、より内側ではある層が脱落して再構成が行われる。海馬体皮質には基本的な多形細胞層、錐体細胞層、分子層よりなる3層構造がある。これらの細胞の軸索、樹状突起の配列によりいくつかの層が二次的にできる。錐体細胞の軸索は海馬白板に投射し脳弓の海馬采となる。)
- 855_31【Collateral eminence of lateral ventricle側副隆起(側脳室の) Eminentia collateralis ventriculi lateralis】 Elevation in the lateral arch of the inferior horn produced by the collateral sulcus.
→(側脳室の側副隆起は海馬近傍で下角外側底において側副溝により生ずる隆起。)
- 855_32【Dentate gyrus; Fascia dentata歯状回 Gyrus dentatus; Fascia dentata】 Convolution between the hippocampal sulcus and fimbriodentate sulcus. It is continuous rostrally with the uncus and caudally with the fasdolar gyrus. Fibers from the neocortex travel through it to Ammon's horn.
→(歯状回は海馬を構成する2つの脳回のうちの1つで、もう一つはAmmon角である。多数の刻み目により歯のようにみえる灰白質からなる弓状の回。小帯回の下方のつづきであり、鈎の内側面に達し、海馬と海馬傍回の間にある。 海馬と同様に歯状回も3層からなる。すなわち分子層、顆粒層、多形細胞層である。これらの層は“U”字あるいは“V”字形となって、開いている部分が海馬采の方を向く。歯状回の分子層は海馬溝の深部で海馬の分子層と連続する。顆粒層は球状や卵形の細胞がぎっしりと並び、そこからでる軸索は多形細胞層を通り抜けて海馬の錐体細胞の樹状突起は主として分子層に入り込んでいる。多形細胞層には錐体細胞の変形や、いわゆる籠細胞など数種類の型が見られる。歯状回から海馬体を越えて出ていく線維はない。海馬体の働きに関係して、組織化学的研究が手掛かりをもたらした。中隔-海馬投射はアセチルコリン分解酵素(AChE)と、コリンアセチル転移酵素(ChAT)陽性の線維を含んでいる。海馬体へのノルアドレナリン性支配は青斑核から起始し、中隔領域を経由して投射する。エンケファリン様免疫反応陽性の軸索は歯状回門から出て、苔状線維として海馬錐体細胞の尖頭樹状突起の近位部に終止する。同じくエンケファリン含有軸索線維の第2グループは、嗅内野皮質の外側部から海馬に「貫通線維」として入ってくる。海馬体の線維連絡を解剖学的に調べてみると、嗅覚性入力をうけていて、その大部分が効果器であることを示唆している。主な求心性線維はそれ自身直接嗅覚線維を受けていない梨状葉の一部である嗅内野から起こっている。嗅内野(28野)からの線維は歯状回と海馬の後部全般に分布する。このうち内側部分から起こる線維はいわゆる海馬白板線維線維を通り、海馬の側脳室に面している側から入る。一方、嗅内野皮質外側部分からの線維は、いわゆる貫通線維を形成して海馬台を通る。これら両者は海馬、歯状回の境界部以外のすべての海馬の区域に分布している。その他、海馬台への入力には海馬采を経由する内側中隔核からの投射線維がある。帯状回から前海馬台と嗅内野皮質への投射は帯状束を経由するが、これは海馬台には入らない。嗅内野皮質は海馬へ投射するので、帯状回皮質からのインパルスは結局海馬に中継されることになる。)
- 855_33【Fimbria of hippocampus海馬采 Fimbria hippocampi】 Fibrous band that leaves the hippocampus and extends as the fornix to the mammillary body.
→(海馬采は白線維質の狭く鋭い縁をした釆状構造で、海馬白板と連続して、海馬の内側縁にある。脳弓脚に続いて前下方に海馬の内側部と癒着しつつ走って海馬傍回の鈎に至る。脳弓は主として海馬、海馬台などから出て海馬白板を通って脳弓に入り、乳頭体などに至る海馬乳頭路からなる。成書によっては脳弓脚よりの海馬采を脳弓采として区別しているものもある。)
- 855_34【Taenia of fimbria采ヒモ Taenia fimbriae】
→()
- 855_35【Hippocampus海馬 Hippocampus】 Crescent-shaped, elongated elevation on the floor of the inferior horn of lateral ventricle. It forms the main part of the archicortex.
→(海馬は(海馬はウマの前半身と長い魚の尾をもったギリシャ神話における動物)またはアンモン角(アンモンはエジプトの巻いた角をもった神)は海馬溝による長い弯曲したたかまりで、側脳室下角の内側壁の大部分を占め、その肥厚した前端部の上面にはいくつかの指のような肥厚があり、これは海馬足または海馬指とよばれ、ほとんど下角の前端に達している。海馬の側脳室表面は海馬白板によっておおわれる。これは内側方は海馬采に続く。海馬采は海馬をおおってその内側部に付く。大脳半球内側面の部分から形成される古い皮質(原皮質)で解剖学的に海馬を厳密に定義することはむずかしい。すでに爬虫類で海馬に相当する皮質がみられ、哺乳類でみられる基本的な要素が発達している。ある種の出力と入力が共通しており、互いに密接に関連(結合)している単純な層形成をもっている部分、すなわち、固有の海馬(アンモン角)と歯状回および、ときに海馬台(海馬支脚、海馬床)、海馬采、脳弓を加えて考えた方が研究目的上都合がよいことが多い。また、しばしばこの意味で、海馬体(hippocampal formation)という言葉が用いられる。さらにBrodmannの脳区分27野(海馬支脚前野),29e野(脳梁膨大後野)、49野(海馬支脚傍野),28野(内嗅領),からなるretrohippocampal foramtionを含めて海馬領域(Angevin, 1965)とよばれる。海馬は構造上部位差がみられ、CA1~CA4の亜核に分けられる(Lorente de No, 1933, 1934)。また、構造上、分子層、錐体細胞層および多形細胞層の各層に区別される。海馬からの遠心性線維は、主として前部から中核部に、また、主として後部から脳弓線維となって視床下部とくに乳頭体に終わる。求心性線維には、嗅内野(entorhinal area, 28野)や嗅周野からくる穿通線維および中隔核や対角帯核からくる中隔海馬線維がある。嗅脳からの直接投射はない。海馬は以前、嗅覚系に関係すると考えられ、ついで情動に、最近では臨床的知見により記憶、とくに短期記憶に関係があることが示唆している。側頭葉基底部の皮質および海馬を両側性に除去すると最近のできごとに関係する記憶が失われる。患者はまったく正常に会話についていけるが、話題が変わるとたちまち前の話の筋を忘れてしまう。)
- 855_36【Crus of fornix脳弓脚 Crus fornicis】 Posterior crus of the fornix that is derived from the fimbria of hippocampus. It encircles the pulvinar and joins with the crus from the opposite side to form the body of fornix.
→(脳弓脚は左右がしだいに離れつつ下外側方に走り、ついで側脳室の下角に向かう。脳梁の下方で左右の脳弓脚の間には脳弓交連がある。)
- 855_37【Collateral trigone側副三角 Trigonum collaterale】 Widened beginning of the collateral eminence at the border of the posterior horn.
→(側副三角は脳梁の膨大部に近い部分で、側脳室の体部が下角および後角と合流する部位を形成している。)
- 855_38【Occipital horn of lateral ventricle; Posterior horn of lateral ventricle後角;後頭角(側脳室の) Cornu occipitale ventriculi lateralis; Cornu posterius ventriculi lateralis】 It extends into the occipital lobe.
→(側脳室の後角(後頭角)は側頭葉中に向かって突出し、その上壁および外側壁はやはり壁板からなる。その他の壁は後頭葉の髄室によって作られ、内側壁には上下各1個のかたまりがある。上のものは不定で、後角球と呼ばれ、脳梁膨大から後方に延びる線維群、すなわち大鉗子によって作られる。下のものは恒存し、鳥距溝のため生じたもので、鳥距という。)
- 855_39【Habenular trigone手綱三角 Trigonum habenulae】 Widening of the habenula at its junction with the thalamus. Location of the two nuclei.
→(手綱三角は手綱の外側端は三角状に広がっており、その吻側より視床髄条が入る。手綱三角の深部には手綱核が存在する。)
- 855_40【Habenular commissure手綱交連;松果交連 Commissura habenularum; Commissura pinealis】
→(手綱交連は左右の手綱核からの線維の交叉。交叉部は松果陥凹の上方にある。)
- 855_41【Occipital lobe後頭葉 Lobus occipitalis】 Lobe that is partially bounded by the transverse occipital sulcus, parieto-occipital sulcus, and preoccipital notch.
→(後頭葉は大脳半球の後部に位置し、外側面では頭頂葉および側頭葉の後方に続き、上外側面ではこれらとは明瞭な境界はない。しかし大脳半球内側面では頭頂後頭溝により頭頂葉と明確に区分される。機能的には視覚野がある。頭頂間溝の後端にほぼ横に走る小溝、すなわち横後頭溝がある。上外側面における溝および回は一般に不規則で、これらを上および外側後頭溝ならびに、上および外側後頭回と呼ぶ。外側後頭溝の後部はしばしば前方に凸部を向けた弓状を呈し、前部の溝と叉状に交わる。この弓状の溝は月状溝または猿裂と呼ばれる。しかしサルの月状溝はヒトの月状溝、頭頂後頭溝および横後頭溝の連続したものと考えられる。)