Spalteholz HANDATLAS DER ANATOMIE DES MENSCHEN VON WERNER SPALTEHOLZ
メニューは解剖学(TA)にリンクしてあります。図の番号をクリックすると下記の説明へ、右側の用語をクリックすると解剖学(TA)にジャンプします。
895


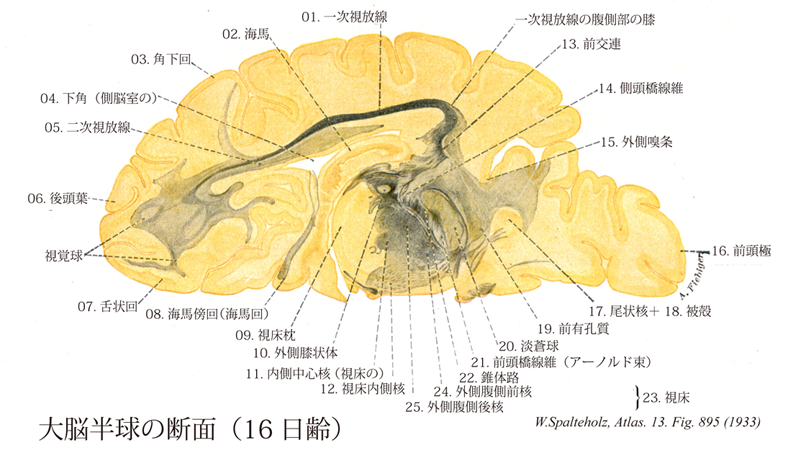
- 895_01【Primary optic radiation一次視放線 Radiatio optica primarius】
→(膝状体皮質路+視床枕皮質路)
- 895_02【Hippocampus海馬 Hippocampus】 Crescent-shaped, elongated elevation on the floor of the inferior horn of lateral ventricle. It forms the main part of the archicortex.
→(海馬は(海馬はウマの前半身と長い魚の尾をもったギリシャ神話における動物)またはアンモン角(アンモンはエジプトの巻いた角をもった神)は海馬溝による長い弯曲したたかまりで、側脳室下角の内側壁の大部分を占め、その肥厚した前端部の上面にはいくつかの指のような肥厚があり、これは海馬足または海馬指とよばれ、ほとんど下角の前端に達している。海馬の側脳室表面は海馬白板によっておおわれる。これは内側方は海馬采に続く。海馬采は海馬をおおってその内側部に付く。大脳半球内側面の部分から形成される古い皮質(原皮質)で解剖学的に海馬を厳密に定義することはむずかしい。すでに爬虫類で海馬に相当する皮質がみられ、哺乳類でみられる基本的な要素が発達している。ある種の出力と入力が共通しており、互いに密接に関連(結合)している単純な層形成をもっている部分、すなわち、固有の海馬(アンモン角)と歯状回および、ときに海馬台(海馬支脚、海馬床)、海馬采、脳弓を加えて考えた方が研究目的上都合がよいことが多い。また、しばしばこの意味で、海馬体(hippocampal formation)という言葉が用いられる。さらにBrodmannの脳区分27野(海馬支脚前野),29e野(脳梁膨大後野)、49野(海馬支脚傍野),28野(内嗅領),からなるretrohippocampal foramtionを含めて海馬領域(Angevin, 1965)とよばれる。海馬は構造上部位差がみられ、CA1~CA4の亜核に分けられる(Lorente de No, 1933, 1934)。また、構造上、分子層、錐体細胞層および多形細胞層の各層に区別される。海馬からの遠心性線維は、主として前部から中核部に、また、主として後部から脳弓線維となって視床下部とくに乳頭体に終わる。求心性線維には、嗅内野(entorhinal area, 28野)や嗅周野からくる穿通線維および中隔核や対角帯核からくる中隔海馬線維がある。嗅脳からの直接投射はない。海馬は以前、嗅覚系に関係すると考えられ、ついで情動に、最近では臨床的知見により記憶、とくに短期記憶に関係があることが示唆している。側頭葉基底部の皮質および海馬を両側性に除去すると最近のできごとに関係する記憶が失われる。患者はまったく正常に会話についていけるが、話題が変わるとたちまち前の話の筋を忘れてしまう。)
- 895_03【Subangular gyrus角下回 Gyrus subangularis】
→()
- 895_04【Inferior horn of lateral ventricle; Temporal horn of lateral ventricle;下角;側頭角;側頭部(側脳室の) Cornu temporale ventriculi lateralis; Cornu inferius ventriculi lateralis】 It extends with the hippocampus laterally and contains the choroid plexus.
→(側脳室の下角(側頭角)は側頭葉に向かって前下方に突出した部分で、この上外側壁は主として脳梁膨大から放散する脳梁線維からなり、これを壁板という。下壁には側副溝によるたかまり、すなわち側副隆起があり、その後方は三角形をなし、側副三角と呼ばれ、後角まで延びている。内側壁は複雑で、上壁との境界部に尾状核尾および分界条があり、その下に上衣層によっておおわれた側脳室脈絡叢があり、さらにその下には脳弓脚の続きである海馬采がある。)
- 895_05【Secondary optic radiation二次視放線 Radiatio optica secundarius】
→(皮質視蓋路)
- 895_06【Occipital lobe後頭葉 Lobus occipitalis】 Lobe that is partially bounded by the transverse occipital sulcus, parieto-occipital sulcus, and preoccipital notch.
→(後頭葉は大脳半球の後部に位置し、外側面では頭頂葉および側頭葉の後方に続き、上外側面ではこれらとは明瞭な境界はない。しかし大脳半球内側面では頭頂後頭溝により頭頂葉と明確に区分される。機能的には視覚野がある。頭頂間溝の後端にほぼ横に走る小溝、すなわち横後頭溝がある。上外側面における溝および回は一般に不規則で、これらを上および外側後頭溝ならびに、上および外側後頭回と呼ぶ。外側後頭溝の後部はしばしば前方に凸部を向けた弓状を呈し、前部の溝と叉状に交わる。この弓状の溝は月状溝または猿裂と呼ばれる。しかしサルの月状溝はヒトの月状溝、頭頂後頭溝および横後頭溝の連続したものと考えられる。)
- 895_07【Lingual gyrus舌状回 Gyrus lingualis】 Continuation of the parahippocampal gyrus to the occipital lobe.
→(紡錘状回と舌状回をそれぞれ外側後頭側頭回、内側後頭側頭回としている成書もあるが、ここでは紡錘状回を内側後頭側頭回とし、舌状回を海馬傍回の後頭方向へのつづきと定義する。つまり、外側後頭側頭回は側頭葉下縁で切れ目なく下側頭回へ移行することになる。舌状回は大脳の内側で側頭葉と後頭葉の境界領域で、水平に走る比較的短い回。深い側副溝により紡錘状回 fusiform gyrusから区切られ、また鳥距溝により楔部から分けられている。その前端は海馬傍回の峡に隣接する。鳥距溝の下縁を形成する回の虫部と上部の帯は、有線領(一次視覚領)の下半分に対応し、両眼視の際、視覚野の反対側の上部4分の1を占める。)
- 895_08【Parahippocampal gyrus; Hippocampal gyrus海馬傍回;海馬回 Gyrus parahippocampalis; Gyrus hippocampi】 Convolution situated below the hippocampal sulcus.
→(海馬傍回は海馬溝の下方、側副溝との間に大きな回である。海馬傍回の皮質の細胞構築は海馬台前部から順次海馬台、前海馬台を経て海馬体、歯状回にいたる間に次第に6層から3層構造に移り変わる。嗅内領(28野)は6層構造の皮質であるが、より内側ではある層が脱落して再構成が行われる。海馬体皮質には基本的な多形細胞層、錐体細胞層、分子層よりなる3層構造がある。これらの細胞の軸索、樹状突起の配列によりいくつかの層が二次的にできる。錐体細胞の軸索は海馬白板に投射し脳弓の海馬采となる。)
- 895_09【Pulvinar of thalamus視床枕 Pulvinar thalami】 Posterior, freely projecting portion of the thalamus.
→(視床枕は視床の後部と背外側を形成する大きな灰白質塊で、これの尾方は、内側膝状体、外側膝状体および中脳の背外側面の上に張り出している。視床枕は視床枕核群ともよばれており細胞学的にはかなり均一であるので、局所的な位置関係を基にして細分される。視床枕を形成するのは、明るく染まった、中等度の大きさの、多極性の細胞で、それらの細胞の分布密度と配列は、視床枕の部位によって異なっている。視床枕の前部の細胞は小さく、明るく染まり、散在性に配列している。下部は、視床枕の主部から、上丘腕の神経線維によって隔てられており、散在性の濃染する細胞で構成される。外側部を、外側髄板から広がる斜めの線維束が横切っている。視床枕の諸核は、長い、上行性の感覚神経路からは入力を受けてはいないが、その下部は、上丘の浅層のいくつかの層からの投射を受けている。局所的には、この投射は、反対側の視野の半分に相当する。視床枕の下部とそれい隣接する外側部とは、線条野を含む後頭葉の皮質と相互に結合する。視床枕の下部とそれに隣接する外側部はそれぞれ、反対側の視覚野の半分が復元され、網膜の部位局在的に、次の各部に投射している。すなわち、①皮質の18野と19野と、②有線野(17野)で、そこでは、線維が、顆粒層の上にある諸層に終止する。これらの結果によって、3つの視覚局在性をもった系統の入力が、視床(外側膝状体、視床枕の下部、およびそれに隣接する視床枕の外側部)から、一次視覚野に達し、しかも、そのいろいろな層に終止することが明らかになった。視床枕の下部から17野、18野および19野への投射は、膝状体外視覚神経路の中の最後の連絡を形成する。(視床枕の下核に隣接している部分以外の)視床枕の外側核は、側頭葉に投射し、同じ領域と相互に連絡している。視床枕の内側部は、上側頭回に投射しているらしい。)
- 895_10【Lateral geniculate body外側膝状体 Corpus geniculatum laterale】 Part of the visual pathway that is connected with the superior colliculus and visual cortex.
→(外側膝状体は、視床の後下面よりわずかに突出している1対の小さな卵形の塊の外側部分。視覚系における視床の中継核であり、内側膝状体の吻側外側方、大脳脚の外側方で、視床枕の副側方にある。この核には、細胞が層状に配列した構造があり、横断切片では、門を腹内側方に向けた馬蹄形をしている。視索の視交叉および非交叉性線維は、この門を通ってはいり、一定の正確な様式に従って分布する。ヒトと霊長類では、外側膝状体を構成するものは、6つの細胞層であり、これらは2つに大別される。同心円状に並んだ6つの細胞層は、介在する線維帯によって区切られ、慣例では、腹内側方の門の領域から始めて、1から6まで番号が付けられている。外側膝状体の背側核を区分すると、大細胞性部(1と2層)および小細胞性部(3~6層)となる。外側膝状核の背側部の2つの領域は網膜の神経節細胞から求心線維を受ける。外側膝状体の小細胞性の層を構成するのは、腹背の方向に順に3,4,5および6層であり、容易に区別される。これらの諸層を外側方にたどると、4層が6層と、また、3層が5層というように対をなす2層が1組になって癒合する。網膜から外側膝状体への投射は正確で、視索の交叉性線維と非交叉性線維とはそれぞれ、別々の層に終わる。すなわち、交叉性線維は1,4および6層に終わり、一方、非交叉性線維は2,3および5層に終わる。交叉性網膜膝状体線維関係のある2つの特殊な性状が構造に反映されている。単眼性の視野は半月形であるが、これは反対側の網膜の内側半の最も内部にある受容要素によって受け取られる。網膜のこの部分にある神経節細胞は、反対側の外側膝状体の二重層の部分に交叉性に線維を送る。この二重層は、4層と6層の部分が外側方で癒合して出来ている。網膜の内側半の中にあって、視神経線維が通っている視神経円板(乳頭)には光受容器がなく、[周辺]視野測定[法]で見つけることができる盲点の原因となる。視神経円板(乳頭)は反対側の外側膝状体の中で、4層と6層の中の細胞層が不連続である部位に対応する。ヒトの外側膝状体を通る切断切片のニッスル標本によって、細胞の直線的配列が明らかにされており、細胞の長軸の方向は、各細胞層の軸に垂直である。小細胞性の諸層にある各周囲部は、“投射の方向線”に平行であり、これらの視野中の各点が同じように復元されていることを示している。外側膝状体の中における網膜表面との局所的対応関係は、高度に組織化され、しかも正確である。両眼視による視野の反対側半は、交叉性線維と非交叉性線維とが異なる層に終わってはいるが、外側膝状体のすべての層に投影される。6つの層における投射の場所は、完全にきまっているので、両眼視における反対側の眼の視野の中のどんなに小さな領域でも、6つの層全体を通して、“投射の方向線”に平行して放射状に延びる背腹方向の細胞柱に一致することを示すことができる。外側膝状体を構成するものは、馬蹄形に曲がった6つの薄い細胞層である。しかし、そこに投射してくる場所は正確にきまっている。それゆえ、“投射の方向線”の中の細胞柱は、両眼視の視野のうち反対側の視野に関係する各眼の網膜の中の対応する点からの入力を受ける。両眼視の像の融合は、外側膝状体の中では起こらない。それは、網膜膝状体線維が外側膝状体の異なった層に終わっているからである。(術後長期間生存させた例の)視神経の切片を追って調べると、順応性の変性、またはニューロン越えの変性は、各側の外側膝状体の3つの層に起こる。神経線維、または細胞の変性が起こる層は、網膜からの交叉性線維(1,4および6層)と非交叉性線維(2,3および5層)の配列に従って異なる。網膜の小さい傷によって、同側、対側の3つの違った層の中に“投射の方向線”に従って、一列に並んで配列した細胞集団に、ニューロン越えの変性が起こる。反対側の単眼視野(単眼性の半月形の視野)は、網膜の内側半のもっとも内側の部分における受容器要素と関係しており、これからは、2層性の部分に終止する交叉性の神経線維のみが出ている。外側膝状体における“投射の方向線”は、有線野に加えた傷害によって外側膝状体の中に現れる逆行性細胞変性の研究からも、明らかにすることができる。視野の半分は、局在性をもってそれぞれの側の半球の有線野に部位局在的に投射しているので、外側膝状体の中に逆行性細胞変性が現れる帯状の部分は、それぞれの側で、投射の方向線によって境される。外側膝状体では視野の中心を通る水平線が背腹方向に入る斜めの面に対応し、この面により内側部と外側部が分けられる。両眼の網膜の上半部からの神経線維は、外側膝状体の内側半部に投射し、下半分は、本核の外側半部に線維を送る。網膜の黄斑からの投射は、外側膝状体尾側部のうち視野の中心を水平子午線に相当する面の両側にある。幅の広いクサビ形の部分として示される。黄斑に相当する部分は、外側膝状体の全容積のおよそ12%を占める。視野の中心を通る垂直の子午線に一致し、本核の尾側縁に沿って、内側の境界からの外側境界に及んでいる。外側膝状体は、視索の主要な終止場所である。ここから膝状体鳥距路、あるいは視放線を経て、鳥距周囲皮質(17野)に投射があり、また外側膝状体は、この皮質野から皮質膝状体線維を受ける。本核は、視床枕と核間結合をしている。)
- 895_11【Central medial nucleus of thalamus内側中心核(視床の) Nucleus centralis medialis thalami; Centre médian de Luys】 Nucleus lying in the lower medial end of the internal medullary lamina. It sends projections mainly to the orbitofrontal and prefrontal cortex.
→(内側中心核は内髄板の下内側端にあり、中心傍核の内側部に隣接して存在する。)
- 895_12【Medial nuclei of thalamus視床内側核群;視床内側核 Nuclei mediales thalami】 Group of nuclei consisting mainly of the dorsomedial nucleus. It receives afferents from other thalamic regions and subcortical structures as well as the amygdaloid body, basal ganglia, and reticular formation of the mesencephalon.
→(視床内側核は視床内髄板と髄板内核に取り囲まれている。その内方および腹内方には視床正中線核群(midline thalamic nuclei)が分布する。正中線核群は菱形核、結合核、および紐傍核である。視床内側核は内側の巨大細胞部(magnocellular part)と外側の小細胞部(paraveocellular part)に区分される。小細胞部と内髄板の境界部に大型細胞から成る髄板傍部(pars paralemellaris)が記載されることがあるが、これはむしろ視床外側中心核(CL)に属するものと考えられる。視床内側核は腹側線条体からの投射もうける。この経路によって「側坐核(腹側線条体のもっとも主要な部分)-視床内側核-前頭前野皮質」の連絡系が成立する。視床から腹側線条体への投射線維は紐傍核(parataenial nucleus)から起こる。)
- 895_13【Anterior commissure前交連 Commissura anterior】 Transverse nerve fibers between the anterior parts of the right and left cerebral hemispheres. It lies posterior to the lamina terminalis.
→(間脳の前交連は第三脳室の前壁をつくる終板の後ろにある横走線維束である。小さくて左右の嗅球を連結する前部と、左右の側頭葉を連結する大きな後部からなる。)
- 895_14Turck, Bundle of【Temporopontine fibres; Temporopontine tract側頭橋線維;側頭橋核線維;側頭橋路;側頭橋核路;側頭部(皮質橋核路の) Fibrae temporopontinae; Tractus temporopontinus】 Fibers passing from the temporal lobe to the pontine nuclei.
→(頭頂、後頭および側頭橋線維で、頭頂葉、後頭葉および側頭葉をなどから出て内方を通って下行し、大脳脚をへて同側の橋核終わる。)
- 895_15【Lateral stria; Lateral olfactory stria外側嗅条 Stria olfactoria lateralis】 Band of fibers lying anterior to the trigone and extending laterally to the insula, then posteriorly to the periamygdaloid cortex.
→(外側嗅条は外側方に走り、島限のあたりで急に曲がって、側頭葉の吻側部の内側部に入る。外側嗅条は、前頭葉の底面を横切って外側方に走る際に、梨状葉前皮質内側部の上面を通る。)
- 895_16【Frontal pole of cerebrum前頭極(大脳の) Polus frontalis】 Anterior end of the frontal lobe.
→(大脳の前頭極は左右大脳半球の最前部の丸く膨隆した部分。)
- 895_17【Caudate nucleus尾状核 Nucleus caudatus】 Elongated nucleus arising from the ganglionic eminence of the telencephalon and curving around the thalamus.
→(尾状核は全体として弓状の大きな灰白質である。尾状核の吻側部は視床の吻側に位置し、側脳室前角のなかに隆起してその外側壁をなし、尾状核頭とよばれる。後方で細くなって尾状核尾とよばれ、視床の背外側縁に沿って側脳室の中心部の底面の外側縁を走り、やがて側脳室の弯曲に沿って下方にまがり、側脳室下核の上壁に達し、扁桃体との後端部のレベルでその外側部に接しておわる。尾状核頭と尾状核尾の中間部を尾状核体という。神経細胞には、大きく分けて、大小2種類(1:20)のものがみられる。求心性神経線維の起始部の主なものは、大脳皮質・視床髄板内核・視床正中中心核・黒質である。黒質(とくに緻密部)からはドパミン作働性の神経線維を受ける。尾状核からの遠心性神経線維の主な分布域は淡蒼球と黒質(とくに網様部)であり、これらの多くのものはGABA作働性である。)
- 895_18【Putamen被殻 Putamen】 Lateral telencephalic part of the lentiform nucleus.
→(被殻はレンズ核の外側部を形成し、外側髄板によって淡蒼球の外節とへだてられている。島皮質とは最外包、前障、外包によってわけられる。被殻の構造は尾状核とまったく同様で、太い有髄線維をほとんど含まず、主として小さい神経細胞からなるが、散在性の大細胞を含む。被殻と尾状核は発生学的にみると、同一の細胞群が内包の発達によって隔てられたもので、両者の間には互いに結合する灰白質の線条が多数見られる。そのため、両者をあわせて線条体または新線条体と呼ぶ。線維連絡も尾状核と原則的に等しい。霊長類において動物が高等になると、相対的な意味で尾状核の体積が減少し、被殻の体積が増大するといわれている。)
- 895_19【Anterior perforated substance前有孔質;嗅野;嗅覚野 Substantia perforata anterior; Substantia perforata rostralis; Area olfactoria】 Perforated area between the olfactory striae produced by the passage of cerebral vessels. It transitions into the gray substance of the tuber cinereum and paraterminal gyrus.
→(前有孔質は嗅脳後部の大部分を占め、内側、外側嗅条と視索が嗅回をなしている。視床、大脳核、内包などにいたる小血管が出入りするため多数の孔が明いている。後縁はブローカの対角帯により形成される。)
- 895_20【Pallidum; Globus pallidus; Paleostriatum淡蒼球;古線条体;旧線条体 Pallidum; Globus pallidus】 Structure arising in the diencephalon and forced by the internal capsule away from its original site, with the greater portion of the site giving rise to the subthalamus.
→(淡蒼球はレンズ核の最内側部を占め、被殻よりは小さい灰白質。垂直に走る板状の有髄神経線維(外側髄板)によって外側部の被殻と隔てられており、また、内側髄板によって内節と外節に分かれる。系統発生学的に線条体よりは古く、下等動物でよく発達している。発生に関しては、「間脳」性とするものののほか、その一部を「終脳」由来とするものがある。鉄反応が強陽性にでることが知られている。淡蒼球には大型の紡錘形ニューロンが多く、樹状突起は無棘で、長く髄板に平行して円板状に分枝する。淡蒼球の大型ニューロンの定量分析によれば、これらの細胞は単一のニューロン群に属する。淡蒼球の内節と外節の大型ニューロンには、なんら形態学的又は化学的な差異は見られない。ヒトでは外節は淡蒼球全体の約70%を占め、細胞密度が最も高い。淡蒼球の内節、外節の大型ニューロンはすべてGABA作働性である。淡蒼球ニューロンの軸索は少数の側枝を出している。淡蒼球の内側・外側髄板に存在する大型のコリン作働性ニューロンは、淡蒼球の腹側に存在する無名質のつづきである。多数の有髄線維束が淡蒼球を横走しているので、新鮮な脳では被殻や尾状核に比べ色が淡く見える。淡蒼球への求心性神経線維のもっとも主な起始は尾状核と被殻である。また、視床下核や黒質からの神経線維が内包を横切って主として内節に入る。淡蒼球が大脳皮質からの求心線維を受けるかどうかについては不確かである。視床からの求心線維については否定的な見解が多い。淡蒼球からおこる遠心性神経線維の主なものは視床と視床下核におわる。これらのうち、内節の腹側から出る神経線維は淡蒼球の腹側表面に集合してレンズ核ワナを形成し、内方後脚の腹内側部を背方にまわり、背尾側方へ走ってフォレル野に達する。一方、内節の背側からでる神経線維は淡蒼球の背内側部からおこる。これらは多数の小線維束として内包の腹側部を横切り、不確帯の腹側部に集合してレンズ核束(H2)を形成する。ついで、レンズ核束はフォレル野に入り、ここでレンズ核ワナの神経線維と一つになり、不確帯の内側端を取り囲むように腹側から背側にまわり、視床束(H1)を形成する。視床束は不確帯の背側を吻外方へ走り、視床に入って主として前腹側核(VA)、外腹側核(VL)、正中中心核(CM)などに分布する。なお、視床束には小脳視床線維も含まれている。淡蒼球から視床下核にいたる神経線維は主として外節からおこり、内包の尾側レベルにおいてその腹内側部を横切って視床下核に達する。淡蒼球からおこり中脳被蓋(脚橋被蓋核)や黒質へ向かう遠心性神経線維もある。)
- 895_21Arnold, Tract of【Frontopontine fibres前頭橋線維;前頭橋路;前頭橋核路;前頭部(皮質橋核路の);アーノルド束 Tractus frontopontinus】 Fibers from the frontal lobe to the pontine nuclei.
→(前頭橋線維は前頭葉、特に中心後回より起こる皮質橋伝導路。大脳脚の内側1/6のところにあり、前頭葉と橋を結ぶ。)
- 895_22【Pyramidal tract錐体路 Tractus pyramidalis】 Tract arising from the cerebral cortex, especially from regions in the frontal and parietal lobes. It transmits activating and inhibiting impulses for voluntary motor function.
→(錐体路本来の定義に従えば、起始領域、終枝部位に関係なく延髄の錐体(pyramis)を通るすべての神経線維群をいう。鳥類以下には見られず、哺乳類とくにヒトでよく発達しており意識的運動を司る。これらの大部分の線維は大脳皮質からおこり脊髄におわる皮質脊髄線維(または路)からなるが、若干の線維は錐体の経過中またはそれよりも前方のレベルでの神経路から離れて脳幹にある反対側の運動性の脳神経核および付近の毛様体(皮質網様体線維)におわる。これらの皮質核線維とよばれるものは厳密には錐体路に含まれないが、しばしば両者(皮質脊髄線維と皮質核線維)を一緒にして錐体路とよばれる。錐体路の起始細胞は、昔からの考えによれば起始細胞は、運動領皮質(4野)の第5層の巨大錐体細胞(Betz)で、その経路は、終脳の内包、中脳の大脳脚、橋の橋縦束、さらに延髄の錐体を下行し、脊髄前角にいたる有髄線維の集まりの長下行路である。その経路中、橋核、脳幹の網様体や運動核、またおそらく大脳基底核などに一部側枝を与え、延髄下端で大部分(91~97%)の線維が交叉し(これを錐体交叉という)これらは対側の脊髄側索(錐体側索路、外側皮質脊髄路)を下るが、小部分はそのまま同側の前索(錐体前索路、前皮質脊髄路)を下行する。しかし、錐体路の大脳皮質の起始領野をみれば、運動領(4野)のみでなく知覚領や連合領を含む他の領野まで包括される。起始ニューロンもBetzの巨大細胞のみならず、第5層に、みられる中型・小型の錐体細胞も証明されている。さらに脊髄の終枝部位についても前角の運動ニューロンに直接おわるものは動物による実験的研究で判明した限りではむしろ少なく、大部分は中間帯や後核基部におわり、介在ニューロンを介して運動ニューロンに影響を与えると思われる(間接皮質運動路)。錐体路の起始・終枝の問題だけでなく、錐体を構成する軸索には、古典的な錐体路以外の錐体外路系の線維も少量ながら含まれており、厳密には、延髄の錐体を通る線維群(錐体路)とそれ以外の運動系(錐体外路)とに分けることはむずかしい。)
- 895_23【Thalamus; Dorsal thalamus視床;背側視床 Thalamus】 Part extending from the interventricular foramen to the tectal plate. Medially it borders on the third ventricle, laterally on the internal capsule and basal ganglia. It is formed by a collection of nuclei derived during ontogenetic development from the dorsal thalamus.
→(視床は、間脳の大きいほうの背側部分を形成する灰白質。背側間脳溝と視床下溝の間の領域であるが、発生の間に大きく発育して、間脳背側部の広い範囲を占めるようになる。間脳は個体発生上、背側視床、腹側視床、視床下部および視床上部の四つの部位に分けられるが、その中で最も大きな部位を占めるのが背側視床である。単に視床といった場合は背側視床を指す。視床は第三脳室の両壁をなす卵円形の構造で、背側の遊離面は薄い髄質から成る帯層におおわれ、肺内側端に視床上部の構造である視床髄条が、前端より後方に走り手網核に付く。また背外側端は分界条によって終脳の尾状核と、外側方は外髄板によっておおわれ腹側視床の視床網様核と境されている。左右の視床は第三脳室内にまたがる視床間橋(中間質)によってつながり、視床下溝で視床下部と境される。視床の内部を構成している視床核は視床脚を介して大脳皮質と相互に結合する。内部には内髄板とよばれる線維板視床を内側部、外側部および前部に分けている。視床は感覚系と統合系との非常に重要な連絡部位である。嗅覚路以外のすべての感覚路がそれぞれ相当する視床の領域に投射する。「最近の研究によれば、嗅覚系も視床を投射する可能性がある」。視床で処理された感覚系情報の流れは視床大脳皮質線維を経て大脳皮質へと送られるが、大脳皮質の側からは多数の大脳皮質視床線維を介して視床における情報処理系に影響が及んでおり、したがって、視床と大脳皮質とは一つの機能単位としてはたらく。「運動」情報は小脳と大脳基底核を経て伝達され、統合系(大脳辺縁系や脳幹網様体など)からのさらに複雑な情報も視床に達する。したがって、視床は一方では大脳辺縁系と脳幹網様体との連結点として機能し、他方では大脳皮質も連絡しているわけである。)
- 895_24【Anterior ventrolateral nucleus of thalamus外側腹側核の吻側部;外側腹側前核(視床の) Nucleus anterior ventrolateralis thalami】 Anterior and lateral portion of the nucleus. It receives afferent fibers from the globus pallidus and sends projections to the premotor cortex.
→(外側腹側核群(VL)に属する。)
- 895_25【Posterior ventrolateral nucleus of thalamus外側腹側核の尾側部;外側腹側後核(視床の) Nucleus posterior ventrolateralis thalami】 Posterior and medial portion of the nucleus. It receives afferents from the cerebellum and has reciprocal connections with the motor cortex.
→(外側腹側核群(VL核)に属する。)