Spalteholz HANDATLAS DER ANATOMIE DES MENSCHEN VON WERNER SPALTEHOLZ
メニューは解剖学(TA)にリンクしてあります。図の番号をクリックすると下記の説明へ、右側の用語をクリックすると解剖学(TA)にジャンプします。
871


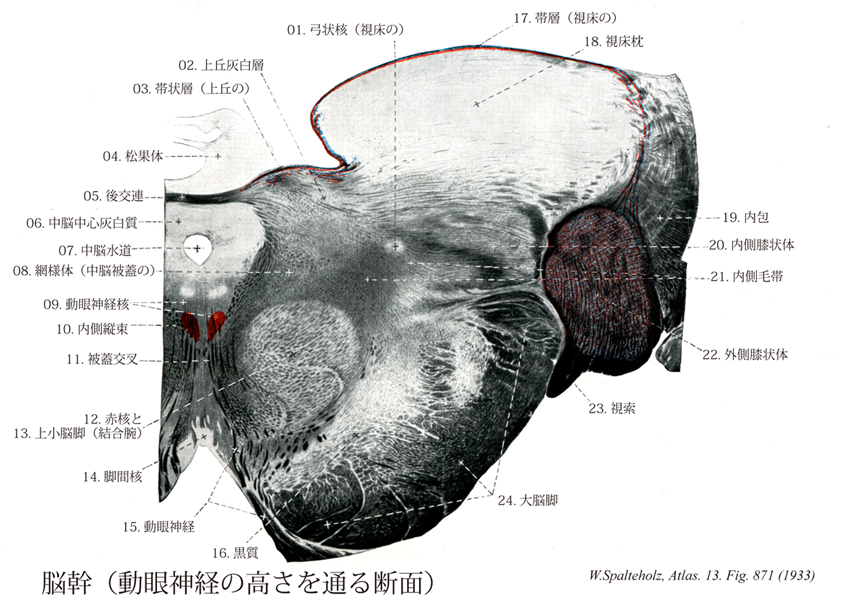
- 871_00【Brain stem脳幹 Truncus encephali】 Collective anatomical term for the rhombencephalon and mesencephalon. The clinical definition includes the basal ganglia, diencephalon, and portions of the 【rhinencephalon】.
→(TAで脳幹は髄脳(延髄)、橋、中脳を脳幹と定義している。かつては、脳を運ぶ際に脳幹をもって運んでいた、脳幹は、脳の柄のような部分という意味で付けられた。日本の神経解剖の教科書では間脳を含める場合や間脳と大脳核を含めて脳幹と称している場合が多いので注意をようする。)
- 871_01【Ventral posteromedial nucleus of thalamus; VPM後内側腹側核;半月核;弓状核(視床の) Nucleus ventralis posteromedialis thalami; Nucleus arcuatus thalami】 Nucleus lying between the ventral posterolateral nucleus and the centromedian nucleus. It receives fibers from the trigeminal lemniscus.
→(VPM核とも言われる。後内側腹側核は比較的淡く染まる神経網からなる三日月形の核で、後外側腹側核の内側で正中中心核の弯曲する境界の外側に位置する。後内側腹側核は次の明確な部分からなる。①小型、大型からなる主部で単に後内側腹側核とよばれる。②弓形の核の内側尖を占める小細胞性で淡染色の部で、小細胞部(VPMpc)とよばれる。後内側腹側核の主部は頭部、顔面、口腔内構造の受容期から体性求心性線維を受け、後外側腹側核の小細胞部は味覚に関係する。後内側腹側核の正確な境界はその線維結合に基づいて規定される。後内側腹側核は上行性二次賛辞神経線維を受けるがこれには、①内側毛帯に伴って上行する三叉神経脊髄路核と主感覚核からの交叉性線維、②三叉神経主感覚核の背側部からおこる背側三叉神経路の非交叉性線維が含まれる。ネコとサルの後内側腹側核からのHRP逆向性輸送により、対側の三叉神経核、特に中間部の尾側半、および三叉神経主核の腹側部の細胞が標識される。同側性に標識される細胞は大部分背側三叉神経路を生ずる主核の背内側部に限局する。顔面筋の伸展受容期からからのインパルスの一部も後内側腹側核の一部に両側性に伝達される。味覚を伝達する中枢経路はまだよくわかっていないが、味覚に関する感覚インパルスが後内側腹側核の内側小細胞部(VPMpc)に表現されることは確実である。鼓索、舌咽神経の電気刺激により、比較的短い潜時でVPMpcに誘発電位が示され体表面の触刺激にVPMpcは反応しないようである。ラットの味覚の表現は両側性といわれるが、ネコとサルではおもに同側性である。BenjaminとAlbert(Benjamin, R.M. 1959. Cortical and thalamic areas involved in taste discrimination in the albino rat. J. Comp. Neurol. 111: 231-260)は、ラットの皮質味覚野の切除によりVPMpcに限局した視床ニューロンの逆行性変性生じることを示した。動物実験の後内側腹側核の内側部の障害により味覚が失われる。ラットの脳神経の一次求心性線維により受容される味覚インパルスは、孤束核の吻側部に達し、そこで二次”橋味覚野”へ同側性に中継される。ラットの橋味覚野は、三叉神経中脳路と上小脳脚あいだの小細胞性の楔上部にある。この領域の細胞は上小脳脚の両側にあり、通常結合腕傍核とよばれる。これらの核の障害による変性は両側性に背内側被蓋を経てVPMpcへ追跡される。腹側前脳部へ達するといわれるこの橋味覚野からの側副路は、小窩講堂に影響する神経領域への直接の結合と考えられる。サルで迷走神経の入る部位より吻側の孤束核に注入されたアイソトープは、同側の中心被蓋路を経てVPMpcに直接達する上行線維束を標識する3H標識アミノ酸は、結合腕傍核とVPMpcに達する同側性線維束へ追跡された。VPMpcへのHRP注入により逆行性に、孤束核吻側部の外側亜分画のみが標識された。これらの所見は味覚が視床へ伝達される経路の理解を助ける。)
- 871_02【Gray layer of superior colliculus上丘灰白層 Stratum griseum colliculi superioris】
→()
- 871_03【Zonal layer of superior colliculus; Layer I of superior colliculus帯状層;帯層;第Ⅰ層(上丘の) Stratum zonale colliculi superioris; Lamina colliculi superioris I】 Most superficial layer composed mainly of glial fibers.
→(上丘の表面にある白質層で大脳皮質から来た線維のごく一部が終わる。)
- 871_04【Pineal gland; Pineal body松果体;松果腺 Glandula pinealis; Epiphysis cerebri; Corpus pineale】 It develops from the roof of the diencephalon and overlies the tectal plate.
→(松果体は円錐形の小体で、後交連の領域で、第三脳室の天井に付着している。これは痕跡的な腺であるらしく、血管の豊富な結合組織の小柱の網工からなり、その網眼の中には神経膠細胞および松果体細胞pinealocytesあるいはepiphysial cellがみられる。サルの松果体細胞はセロトニン(5-HT)とコレシストキニン(CCK)を含んでいる。哺乳類の松果体細胞は、系統発生学的には、感覚神経性光受容体の要素と関係しており、これは、分泌細胞になる物が多いが、間接的に光受容性をもっている。これらの突起の棍棒状をした終末は、血管血管を取り囲む血管周囲腔に接して終わる。松果体の分泌のうちで最もよく知られた物は、セロトニン、ノルアドレナリンおよびメラトニンという生体産生アミンであるが、その他に、サイロトロピン遊離ホルモン(TRH)、黄体形成ホルモン遊離ホルモン(LHRH)およびソマトスタチン(SRIF)のような、視床下部で形成されると同定されたペプチドを、かなりの濃度で含んでいる。セロトニンは、松果体細胞の中で合成されて細胞外の隙間に放出される。ノルアドレナリンは、松果体の実質細胞に終止ししている交感神経ニューロンの中で合成される。松果体は、昼間光の変動に敏感なN-アセチルトランスフェラーゼおよびヒドロキシインドール-o-メチルトランスフェラーゼという2つの酵素の働きによって、セロトニンからメラトニンを合成する。メラトニン合成の毎日の変動は周期性であり、光入力の毎日の周期に直接関係している。光は、日周期を環境の光周期に一致させる。そして、さらに、まだ確認されていない経路を経て、神経性信号が松果体に運ばれることを速やかに遮断するように働く。N-アセチルトランスフェラーゼの活性は、昼夜に高められるが、光にさらすと、酵素の活性が失われる。視床下部の視交叉上核を両側性に傷害すると、この核は網膜視床下部経路を受けているので、松果体のN-アセチルトランスフェラーゼにおける周期がなくなり、その結果、ヒドロキシインドール-o-メチルトランスフェラーゼの活性のレベルが下がる。そのような傷害によって、自発運動の活性の日周期および栄養補給と水飲み行動の両方の日周期がなくなる。雄のネズミでは、視交叉上核の傷害によって、正常な発情周期がなくなる。網膜視床下部路は、視交叉上核の構造との直接の相互作用によって、松果体のはたらきを変える。環境の光は、日周期をその周期に一致させる働きと伝達する働きをもっているとみなされ得る。内在性振動発生装置を光周期にのせるための光後下はゆっくりであるが、信号伝達に及ぼす光の後下は速やかである。そして、それによって、光によってN-アセチルトランスフェラーゼが速やかに“消失”することと、持続光によって日周期が妨げられることを、おそらくは説明できるであろう。視交叉上核からの単一神経路が、松果体によるメラトニン形成に関係する両酵素を調節しているが、これらの結合についての詳細は、なお、あきらかになっていない。間接的に証明された根拠は視交叉上核から松果体への神経路には、視床下部の灰白隆起の領域、内側前脳束および中間質外側路への中継路が含まれることを示唆している。このように、松果体は、交感神経ニューロンを経て受けた信号をメラトニンという内分泌物に変換させる、神経内分泌変換体ともいうべきものであるらしい。松果体の分泌は視床下部の働きを変化さえるが、それは、内分泌が心身の血液循環あるいは、脳脊髄液に入ってからののちに作用する。松果体のセロトニンとメラトニンの日内変動は、光入力の周期に応じて起こる。松果体活動のこれらの周期性変化は、この腺が生物学的時計としてはたらいて、生理学的過程と行動学的過程とを調節する信号を出していることを示唆している。これらの変動は日周期circadian rhythmsと呼ばれ、環境からの刺激が存在すれば、ほぼ24時間周期を示すだけである。松果体実質の腫瘍は、性的機能を低下させて思春期を送らせるが、他方、松果体を破壊するとしばしば早発思春期を伴う。これらの観察は、松果体が、性腺および生殖器系に抑制的影響を及ぼすことを示す実験的研究の結果と一致する。)
- 871_05【Posterior commissure後交連 Commissura posterior; Commissura epithalamica】 Commissure situated between the pineal recess and the opening of cerebral aqueduct. Crossing site of fibers from the surrounding area.
→(後交連は、中脳と間脳の背側における境界をなす。この小さな交連は中脳水道と第三脳室の移行部で上丘の上方の中心灰白質の背側にある。交連線維は外側で扇状にひろがり、その周囲を後交連核と総称される細胞が取り囲む。これまで明らかにされた後交連の構成線維は、①視蓋前核、②後交連核、③間質核からの線維がある。瞳孔の対光反射に関与する線維は後交連で交叉する。後交連の下方の中脳水道上衣は線毛をもつ高い柱状細胞よりなる。このような変形上衣細胞は交連下器官を形成し、分泌機能をもち、脳室周囲器官の一つとされている。交連下器官は中脳において脳血液関門をもたない唯一の部分である。)
- 871_06【Periaqueductal grey substance; Central grey substance中脳中心灰白質;中脳水道周囲灰白質 Substantia grisea centralis; Stratum griseum centrale】 Gray substance surrounding the cerebral aqueduct.
→(中脳中心灰白質は中脳水道を取り囲み密集する比較的小型の細胞からなり、機能的には均一ではない。この領域はこれまで中枢の鎮痛機構、発生、生殖行動の制御、攻撃行動、上方注視機構と深く関係するとされてきた。この中脳中心灰白質は視床下部、脳幹毛様体、縫線核、青斑核および脊髄からの入力を受け、これらの多くはさらに中脳中心灰白質からの相互の投射を受ける。中脳中心灰白質のニューロンはエンケファリン、P物質、コレシストキニン、ニューロテンシン、セロトニン、ダイノルフィン、ソマトスタチンに対して免疫反応陽性であり、ひとつのニューロンがしばしば複数のニューロペプチドを有する。中脳中心灰白質の腹外側部は、刺激による鎮痛に関して最も効果的な部位と思われる。中脳中心灰白質の腹側部へのモルヒネの微量注入により著明な鎮痛を引き起こすことが齧歯類動物において認められている。)
- 871_07Sylvius, Aqueduct of【Aqueduct of midbrain; Cerebral aqueduct中脳水道;脳水道 Aqueductus mesencephali; Aqueductus cerebri】 Narrow canal in the mesencephalon between the third and fourth ventricles.
→(中脳水道はシルビウス水道ともよばれる。中脳では脳室系は細い管となり、間脳の第三脳室と菱脳の第四脳室とを結合する。これを中脳水道と称し、横断面は円形または底辺を背側に向けた角のとれた三角形をなし、中心灰白質によってかこまれる。その存在については古くから知られていたが、フランスの解剖学者Jacobus Sylvius (1478-1555)の著書(1555年)で初めて説明がなされた。)
- 871_08【Reticular formation網様体(中脳被蓋の) Formatio reticularis tegmentum mesencephali】
→(中脳網様体は橋におけるほど広くない。赤核は網様体の一部ではあるが、通常は網様体といえば赤核を除いてその背側および外側の部分をさす。ここには①楔状核、②楔状下核および③脚橋被蓋核の3つの主な網様核が認められる。)
- 871_09【Nucleus of oculomotor nerve動眼神経核 Nucleus nervi oculomotorii】 Nucleus situated at the level of the superior colliculus anterior to the cerebral aqueduct.
→(動眼神経核は第三脳神経、すなわち、動眼神経の起始核であり、動眼神経運動神経細胞の集合である。中脳被蓋の正中線背側部の両側に存在し、中脳中心灰白質の腹方に位置する。動眼神経核の尾側端は滑車神経核の吻側端とほとんど連続しており、動眼神経核の吻側端は中脳の最吻側レベルに達する。動眼神経運動神経細胞はそれぞれの筋支配に対応して局在配列する。すなわち、下直筋支配細胞は核の背側部、内直筋支配細胞は核の腹側部、下斜筋支配細胞は前2者の中間部、上直筋支配細胞は内側部に位置し、それぞれ脳幹の長軸に平行な左右一対の細胞柱を形成する。これらのうち、上直筋支配細胞柱だけは反対側の筋を支配するが、その他は同側の筋を支配する。また、上眼瞼挙筋を支配する細胞柱は正中部に位置し、不対であって、動眼神経核の尾側1/3のレベルにのみ存在する(尾側正中核)。以上の他、副核(Edinger-Westphal核)および正中核(Perlia核)を含めて動眼神経核群とされることがおおい。)
- 871_10【Medial longitudinal fasciculus; MLF内側縦束 Fasciculus longitudinalis medialis】 Bundle of various fiber systems that enter and leave at different levels. Its fibers interconnect the motor nuclei of cranial nerves and also connect the vestibular apparatus with ocular muscles, neck muscles, and the extrapyramidal system. This serves to coordinate muscle groups, e.g., masticatory, tongue, and pharyngeal muscles when swallowing or speaking; or ocular muscles for movements of the globe.
→(前索の後部には脳幹のいろいろなレベルにある種々な神経核からでる複雑な下行線維束がある。この複雑な神経線維束は内側縦束として知られている。この神経束の脊髄部は同じ名称で呼ばれる脳幹にある伝導路の一部にすぎない。内側縦束の上行線維は主として前庭神経内側核および上核から起こり、同側性および対側性に主として外眼筋支配の神経核(外転、滑車、動眼神経核)に投射する。内側核からの上行線維は主に交叉をし、両側の外転神経核と左右の動眼神経核に非対称性に終わるが、滑車神経核へは対側性に投射する。上核の中心部にある大形細胞は非交叉性上行線維を内側縦束に出し、これは滑車神経核および動眼神経核に終わる。同核の周辺部にある周辺部にある小型細胞は交叉性の腹側被蓋束(内側縦束の外側にある)を経て動眼神経核に投射するが、これは主として対側の上直筋を支配する細胞に作用する。生理学的には、前庭神経核から外眼筋支配核から外眼筋支配核への上行性投射のうち、交叉性線維は促進的に働くが、非交叉性線維は抑制的に働く。内側縦束にはこのほかに、左右の外転神経核の間にある神経細胞から起こり、交叉して上行し、動眼神経核の内側直筋支配部に終わる明瞭な線維が含まれる。この経路は一方の外転神経核の活動を対側の動眼神経核内側直筋支配部へ連絡する物で、外側視の場合に、外側直筋が収縮すると同時に対側の内側直筋が共同して収縮するための神経機構を形成している。内側縦束の上行線維の一部は、動眼神経核を回ってCajal間質核に終わる。これは内側縦束内にうまっている小さい神経細胞群である。前庭神経内側核は対側性に間質核へ投射するが、上核は同側性に終わる。前庭神経二次線維は両側性に視床の中継核へ投射し、その数は中等度で、後外側腹側核に終止する。前庭からの入力を受ける視床の細胞は体性感覚情報にも対応するが、これは視床には特定の前庭感覚中継核がないことを示唆している。)
- 871_11【Tegmental ducussations被蓋交叉 Decussationes tegmentales】 Crossing of tracts in the mesencephalon.
→()
- 871_11aMeynert's decussation【Posterior tegmental decussation; Dorsal tegmental decussation背側被蓋交叉;後被蓋交叉 Decussatio tegmentalis posterior】 Decussation of the tectospinal and tectobulbar tracts. Fibers from the superior colliculus.
→(上丘の深部にある細胞から起こる視蓋脊髄路が、中心灰白質のまわりを前内側方へ走り背側被蓋交叉で交叉する。)
- 871_12【Red nucleus赤核 Nucleus ruber】 Principal nucleus of the central tegmental tract. The iron-containing nucleus is situated between the periaqueductal gray substance and substantia nigra, extending from the superior colliculus into the diencephalon. It is composed of two or possibly more components.
→(赤核は中脳被蓋では最も明瞭な部分で、網様体のほぼ中央にあり、桃黄色を呈し、周囲を上小脳脚線維で囲まれている。赤核全体として卵円形で、横断面は円形を呈し、上丘の下端から間脳の下部まで広がる。細胞学的には後方の大細胞部と前方の小細胞部によりなる。赤核の細胞の間には主として上小脳脚からの有髄線維の細い束が存在する。動眼神経根の一部が脚間窩に至る途中で赤核を貫く。赤核の求心性線維は主として小脳核および大脳皮質からくる。これらの線維は体部的局在性をもって赤核に終止する。上小脳脚の線維は中脳下部で完全に交叉し、対側の赤核およびその周囲に至る。小脳歯状核からの側枝は対側赤核の上1/3部に終止し、一方、栓状核(前中位核)および球状核(後中位核)からの線維は赤核の下2/3部に体部位的局在性をもって終わる。この連絡は小脳の傍虫部皮質と赤核大細胞部をつなぎ、さらに赤核から脊髄へ体部位的局在性に投射する。これには上小脳脚交叉および赤核脊髄路の交叉の2つの交叉が含まれる。赤核の大細胞部からの体部位的局在投射は主に頚髄と腰髄に終わる。皮質赤核線維は中心前回や前運動野から起こり、ともに両側の赤核小細胞部に体部位的局在性をもって終止する。大脳皮質6野の内側部(補足運動野)からの投射線維は対側の赤核大細胞部に終わる。中心前回の運動野から赤核大細胞部に終止する線維は同側性で、赤核運動路の体部位的局在性をもつ起始に対応する。これらの皮質赤核線維は体部位的局在性をもち、赤核脊髄路(交叉性)と共に運動野皮質から脊髄へインパルスを伝える経路の一部をなす。赤核からの下行性遠心路は腹側被蓋交叉で交叉し、①小脳の中位核、②三叉神経主知覚核および脊髄路核、③顔面神経核の一部、④いくつかの延髄の中継核および⑤脊髄に投射する。また赤核の小細胞部からの非交叉性の線維束が中心被蓋路に入り、オリーブ核主核の背側板に終わる。これを赤核オリーブ路といい、小脳へのフィードバック系の一部をなす。赤核からの視床への投射はない。)
- 871_13【Superior cerebellar peduncle上小脳脚;結合腕 Pedunculus cerebellaris superior; Brachium conjunctivum】
→(上小脳脚(結合腕Brachium conjunctivum)は主として小脳を出る線維からなる。その主体をなす線維は小脳視床路と小脳赤核路である。これらは主として歯状核から出て、腹内側方に進んで深部に入り、中脳下半で大部分交叉し、上小脳脚交叉(結合腕交叉)を作り、反対側の中脳被蓋を上行し、一部は赤核に終わるが(小脳赤核路)、一部はさらに視床の前外側腹側核に至る(小脳視床路)。なお上小脳脚の表面を前脊髄小脳路が逆行して小脳に入り、主としてその前葉に分布する。また鈎状束は室頂核から出て大部分交叉し、上小脳脚の背外側をへて鈎状に曲がり、下小脳脚内側部の上部に来て前庭神経各核にならびに橋、延髄の網様体内側部に分布する。)
- 871_14【Interpeduncular nucleus脚間核 Nucleus interpeduncularis】 Cell group overlying the interpeduncular fossa.
→(脚間核は脚間窩の背側にある。この核は手綱核からの投射を反屈束を経由し受けている。多くのコリン作働性線維が脚間核に終止する。反屈束の線維のうちで脚間核を通過するものは上中心核、背側被蓋核および中脳中心灰白質に達している。背側被蓋核はまた乳頭体被蓋路を通して乳頭体からも線維を受け、中脳中心灰白質の腹内側にある小さな線維束である背側縦束と密接に関係している。これらの神経路は辺縁系に関連した信号を中脳の諸核に伝えている複雑な神経路の一部を構成しており、内臓や行動機能に関係する。)
- 871_15【Oculomotor nerve [III]動眼神経[脳神経III] Nervus oculomotorius [III]】 Nerve containing motor and parasympathetic fibers that exits the oculomotor sulcus and passes through the superior orbital fissure into the orbit.
→(動眼神経の主成分は動眼神経主核から出る体性運動性のもので外側直筋および上斜筋以外の眼筋を支配する。このほかに副交感性の動眼神経副核[Edinger-Westphal核]から出る線維が加わる。以上の2核から出る線維は多数の根をつくって大脳脚内側溝から出て1神経幹となり、滑車神経、外転神経および眼神経とともに、蝶形骨体の両側にある海綿静脈洞の上壁に沿ってすすみ、上眼窩裂を通って眼窩内に入り、上下の2枝に分かれる。上枝は上瞼挙筋および上直筋に、下枝は内側直筋、下直筋および下斜筋に分布する。また下枝からはきわめて短い動眼神経からの根が出て、毛様体神経節に入るが、これは動眼神経副核から出て、下枝を通って毛様体神経節に入る副交感線維にほかならない。)
- 871_16【Substantia nigra黒質;黒核 Substantia nigra; Nucleus niger】
→(黒質は中脳被蓋腹側部の核で大脳脚の背側に接して存在する。ヒトの黒質の神経細胞は顆粒状のメラニン色素を豊富に含有するため、黒質は全体として肉眼的に黒くみえる。黒質には背側の緻密部と腹側の網様部が区分される。緻密部が神経細胞に富むのに対し、網様部では神経細胞の密度は粗で、細い神経線維に富む。したがて、前者は黒色部、後者は赤色部とよばれることがある。黒質からおこる遠心性神経線維としては、緻密部からおこり線条体に分布する黒質線条体線維、網様部から起こり視床のとくに内側腹側核(VM)に分布する黒質視床線維、および網様部からおこり上丘の中間灰白質に分布する黒質上丘線維などが主なものである。また、黒質に分布する求心性神経線維の起始としては、線条体・淡蒼球・視床下核(Luys体)が主なものである。これらのほか、前頭葉皮質・背側縫線核・扁桃体中心核・外側手綱核なども報告されているが不確実である。黒質は中枢神経系のうちでドーパミンとGABAの含有量が高い部位として知られる。ドーパミンは線条体に神経線維を送る黒質緻密部の神経細胞に主として含まれ、またGABAは線条体よりおこり黒質網様体に至る神経線維の軸索終末に主として含まれる。黒質に見られる線維はまたは11個のアミノ酸が連絡したペプチドとしてのP物質(SP)も含む。黒質は脳において最も高濃度にP物質を有する部位で、この物質は黒質の緻密部および緻密部内の神経終末に凝集している。網様部はまたエンケファリン作働性線維および終末も有する。尾状核および被殻の樹状突起の棘突起に含むニューロンから起こる線条体黒質線維はGABA、P物質、エンケファリンを含む。これらの線維は同様の伝達物質を有する線条体淡蒼球線維を出すニューロンとは異なる細胞集団から起こる。黒質はパーキンソン病(振戦麻痺)の原因となっている代謝障害に緻密に関係しており、Huntington舞踏病および異常な不随意運動や筋緊張の変化を特徴とする他のタイプの運動障害にも関与しているようである。パーキンソン病では黒質から線条体へのドーパミンの輸送および合成が極度に傷害される。Huntington舞踏病では線条体のドーパミンは正常であるがGABAは著明に減少している。)
- 871_17【Zonal layer of thalamus帯層;帯状層 Stratum zonale thalami】
→()
- 871_18【Pulvinar of thalamus視床枕 Pulvinar thalami】 Posterior, freely projecting portion of the thalamus.
→(視床枕は視床の後部と背外側を形成する大きな灰白質塊で、これの尾方は、内側膝状体、外側膝状体および中脳の背外側面の上に張り出している。視床枕は視床枕核群ともよばれており細胞学的にはかなり均一であるので、局所的な位置関係を基にして細分される。視床枕を形成するのは、明るく染まった、中等度の大きさの、多極性の細胞で、それらの細胞の分布密度と配列は、視床枕の部位によって異なっている。視床枕の前部の細胞は小さく、明るく染まり、散在性に配列している。下部は、視床枕の主部から、上丘腕の神経線維によって隔てられており、散在性の濃染する細胞で構成される。外側部を、外側髄板から広がる斜めの線維束が横切っている。視床枕の諸核は、長い、上行性の感覚神経路からは入力を受けてはいないが、その下部は、上丘の浅層のいくつかの層からの投射を受けている。局所的には、この投射は、反対側の視野の半分に相当する。視床枕の下部とそれい隣接する外側部とは、線条野を含む後頭葉の皮質と相互に結合する。視床枕の下部とそれに隣接する外側部はそれぞれ、反対側の視覚野の半分が復元され、網膜の部位局在的に、次の各部に投射している。すなわち、①皮質の18野と19野と、②有線野(17野)で、そこでは、線維が、顆粒層の上にある諸層に終止する。これらの結果によって、3つの視覚局在性をもった系統の入力が、視床(外側膝状体、視床枕の下部、およびそれに隣接する視床枕の外側部)から、一次視覚野に達し、しかも、そのいろいろな層に終止することが明らかになった。視床枕の下部から17野、18野および19野への投射は、膝状体外視覚神経路の中の最後の連絡を形成する。(視床枕の下核に隣接している部分以外の)視床枕の外側核は、側頭葉に投射し、同じ領域と相互に連絡している。視床枕の内側部は、上側頭回に投射しているらしい。)
- 871_19【Internal capsule内包 Capsula interna; Capsula interna nuclei lentiformis】 Band of nerve fibers lying medial to the lentiform nucleus and medial to the thalamus and caudate nucleus.
→(内包は外側のレンズ核と内側の尾状核および視床との間にある、大きい線維束の集団で、その大部分は下方に集まって大脳脚に移行する。内包は大脳半球の水平断でみると、内包前脚と内方後脚からなり、これらは鈍角をなして交わり、内包膝の名で知られる接合部を形成する。内包前脚はレンズ核と尾状核の間にあり、また内包後脚(レンズ核視床部)はレンズ核と視床の間にある。内包のレンズ後部は尾方に、レンズ核の少し後ろにまで伸びる。この尾方の領域にはレンズ核の下を通って側頭葉に達する一群の線維があり、これらはまとまって内包のレンズ下部を形成する。①視覚、聴覚、体性感覚放線などを構成して視床から大脳皮質へ上行している線維と、②大脳皮質から視床、視床腹側部、中脳、後脳、脊髄へ下降している線維から構成される。)
- 871_20【Medial geniculate body内側膝状体 Corpus geniculatum mediale】 Part of the auditory pathway that is connected with the inferior colliculus.
→(内側膝状体は、視床の尾方腹側面で、外側膝状体の内側方、大脳脚の背方に位置する。この核は、視床における聴覚の中継核であって、聴放線を出す。下位脳幹にある聴覚の中継核群とは違って、両側の内側膝状体の間には交連線維による連絡がない。内側膝状体は、明瞭な細胞構築と連絡から更に幾つかの部分にに分けられる。内側膝状体は内側部、背側部および腹側部と呼ばれる三つの主要な部分から構成される。内側膝状体のこれらの細区分域は、普通の組織学的標本では区別するのが容易ではないが、Golgi標本では明らかである。内側膝状体腹側核は、内側膝状体の吻尾方向の全長にわたって広がり、内方が、下丘腕によって境されている。内側膝状体の他の大きな部分とは違って、腹側核には、明確な層板構造がある。腹側部の細胞の大きさと形は、かなり一定しており、房状の樹状突起をもつ。房状細胞の樹状突起と下丘腕の神経線維によってできた層状構造は、らせん形、または、弯曲した垂直の板状を示す。下丘からの求心性線維は特定の層板にはいり、そのまま同じ層と連絡しつづける。内側膝状体の側腹部にある層板構造は、外側膝状体のそれに類似しているが、細胞の層が有髄神経線維帯によって区切られることがない。内側膝状体の腹側部に生理学的な性質によって地図をつくると、この細胞層は音の高低に一致した局在と関係しており、高い周波数の音は内側に、低い周波数の音は外側に復元される。内側膝状体の腹側部のニューロンから聴放線が起こり、一次聴覚野に終わるが、ここでは、音の周波数が空間的に配置されている。一次聴覚野からは内側膝状体の腹側部に終わる両方向性皮質視床線維が起こる。膝状体皮質線維と皮質膝状体線維は、両者とも同側性である。ヒトでは、内側膝状体の主な皮質投射は、膝状体側頭葉放線、あるいは聴放線を経て、側頭葉上面の隆起部(横側頭回)に達する。皮質のこの投射野(41野)は、音の高低に一致した局在をもっている、すなわち、高音は内側方に、また低音は前外側方に復元される。内側膝状体の背側部には、幾つかの核が含まれ、それらの中には、膝状体上核と背側核がある。背側核は、内側膝状体の尾方の高さでは顕著であり、外側被蓋野からの投射を受ける。この外側被蓋野は上丘の深い層から、外側毛帯に隣接する領域へと広がった領域である。内側膝状体の内側部の大細胞性部は、下丘、外側被蓋および脊髄からの入力を受ける。内側膝状体の中の層構造を示さない部分はすべて一次聴覚野を取り囲む皮質の帯状部に同側性の線維を送っている。)
- 871_21【Medial lemniscus内側毛帯 Lemniscus medialis】 Ascending fibers passing from the decussation of medial lemniscus through the brainstem to the thalamus. They convey impulses of general cutaneous sensation.
→(延髄の後索(薄束および楔状束)を通過する伝導路は圧覚と触覚や固有知覚の興奮を後索核(薄束核および楔状束核)や視床を経て大脳皮質に伝達する神経路である。薄束核および楔状束核から起こる二次ニューロンは延髄視床路(内側毛帯)となり正中縫線近く延髄の中心を通り上行する。橋にはいると外側に広がり、橋核の背側縁を越えて上行する扁平な帯になる。中脳内では、黒質の背側縁を越えて赤核で外側に移る。内側膝状体まで内側を通り視床の後腹側核に入り、そこで終わる。視床を出た第3ニューロンの線維は、上行して大脳皮質におもむく。内側毛帯系は脊髄から上行する識別性感覚路の最初の一環を形成するのは後根を通って入ってくる太い有髄神経の枝であり、後索を上行する。後索を上行するこれらの神経線維は身体部位対応配列を示す。すなわち、仙骨神経根や腰神経塊を通って入ってくる上行枝は後索の内側部を占めて薄束を形成する。一方、頚神経根を通って入ってくる上行枝は後索の外側部を占めて楔状束を形成する。また、胸神経根を通って入ってくる少数の上行枝は、薄束と楔状束との間に位置する。薄束と楔状束は、延髄の尾側端でそれぞれ対応する神経核、すなわち、薄束核と[内側]楔状束核に終止する。ある後根が支配する皮膚領域は、同時にその後根の上下の後根からも支配されている。このように一定の皮膚領域を支配する隣接後根の神経線維群は、後根から後索、さらに後索核へと向かう経過のうちに、一つの神経束にまとまる。このような集束の結果として、隣接する皮節(dermatome)間の重なり合いは解消されるのであるが(一つの皮節からの情報を伝道する神経線維が集合して一つにまとまる)、後索で最初にみられたような層構造は次第に不明瞭になる。薄束核の背側部と[内側]楔状束核の背側部にはニューロンが幾つかの小群をつくって分布する。超す悪を上行する神経線維の中で、四肢の遠位部を支配するものがこれらのニューロン小群に終止して身体部位対応配列を示す。後索核の腹側部と吻側部では身体部位対応配列はあまり精細でない。後索には後核固有核から起こる内在性神経線維も含まれている。これらの内在性神経線維は後索核の腹側部と吻側部に終止する。その他、後側索(側索後部)を上行する神経線維は両側の後索核の腹側部と吻側部、およびZ群(group Z)に終止する。Z群は薄束核の吻側端に位置するニューロン群であり、下肢の筋からの入力を視床に中継する。上枝からの固有感覚性入力を中継するのは外側楔状束核である。この核から起こる投射神経線維は主として下小脳脚を通って小脳へ入る。後索核の腹側部と吻側部から起こる投射神経線維は、後索核背側部(ニューロンの小群の集合から成る)から起こる投射神経線維に比べて、分布範囲が広い。すなわち、前者も後者も反対側の視床へ向かうのであるが、前者はさらに小脳や下オリーブ核に投射し、脊髄の後角へ向かうものもある。薄束核と[内側]楔状束核からは内弓状線維が起こり、正中部で交叉してのち、内側毛帯を形成して上行枝、視床の後外側腹側核、後核群、内側膝状体大細胞部、および不確帯に終止する。後索核から後外側腹側核への投射は”核と殻(core-and-shell)”の様式を示す。すなわち、後索核背側部から起こる皮膚感覚の投射線維は後外側腹側核の中心部(すなわちcoreの部分)に終止し、後索核腹側部と吻側部から起こる固有感覚の投射線維は後外側腹側核の辺縁部(すわなち、shellの部分)に終止する。内側毛帯線維は後外側腹側核において一連の平行な層板をなして終止する。これらの層板は核のcoreの部分とshellの部分を通じて前後方向に伸びており、それぞれの層板が身体の特定の部位に対応している。また、各層板の前後軸に沿って、種々の感覚要素に対応する投射線維の終末が次々と配列分布する。)
- 871_22【Lateral geniculate body外側膝状体 Corpus geniculatum laterale】 Part of the visual pathway that is connected with the superior colliculus and visual cortex.
→(外側膝状体は、視床の後下面よりわずかに突出している1対の小さな卵形の塊の外側部分。視覚系における視床の中継核であり、内側膝状体の吻側外側方、大脳脚の外側方で、視床枕の副側方にある。この核には、細胞が層状に配列した構造があり、横断切片では、門を腹内側方に向けた馬蹄形をしている。視索の視交叉および非交叉性線維は、この門を通ってはいり、一定の正確な様式に従って分布する。ヒトと霊長類では、外側膝状体を構成するものは、6つの細胞層であり、これらは2つに大別される。同心円状に並んだ6つの細胞層は、介在する線維帯によって区切られ、慣例では、腹内側方の門の領域から始めて、1から6まで番号が付けられている。外側膝状体の背側核を区分すると、大細胞性部(1と2層)および小細胞性部(3~6層)となる。外側膝状核の背側部の2つの領域は網膜の神経節細胞から求心線維を受ける。外側膝状体の小細胞性の層を構成するのは、腹背の方向に順に3,4,5および6層であり、容易に区別される。これらの諸層を外側方にたどると、4層が6層と、また、3層が5層というように対をなす2層が1組になって癒合する。網膜から外側膝状体への投射は正確で、視索の交叉性線維と非交叉性線維とはそれぞれ、別々の層に終わる。すなわち、交叉性線維は1,4および6層に終わり、一方、非交叉性線維は2,3および5層に終わる。交叉性網膜膝状体線維関係のある2つの特殊な性状が構造に反映されている。単眼性の視野は半月形であるが、これは反対側の網膜の内側半の最も内部にある受容要素によって受け取られる。網膜のこの部分にある神経節細胞は、反対側の外側膝状体の二重層の部分に交叉性に線維を送る。この二重層は、4層と6層の部分が外側方で癒合して出来ている。網膜の内側半の中にあって、視神経線維が通っている視神経円板(乳頭)には光受容器がなく、[周辺]視野測定[法]で見つけることができる盲点の原因となる。視神経円板(乳頭)は反対側の外側膝状体の中で、4層と6層の中の細胞層が不連続である部位に対応する。ヒトの外側膝状体を通る切断切片のニッスル標本によって、細胞の直線的配列が明らかにされており、細胞の長軸の方向は、各細胞層の軸に垂直である。小細胞性の諸層にある各周囲部は、“投射の方向線”に平行であり、これらの視野中の各点が同じように復元されていることを示している。外側膝状体の中における網膜表面との局所的対応関係は、高度に組織化され、しかも正確である。両眼視による視野の反対側半は、交叉性線維と非交叉性線維とが異なる層に終わってはいるが、外側膝状体のすべての層に投影される。6つの層における投射の場所は、完全にきまっているので、両眼視における反対側の眼の視野の中のどんなに小さな領域でも、6つの層全体を通して、“投射の方向線”に平行して放射状に延びる背腹方向の細胞柱に一致することを示すことができる。外側膝状体を構成するものは、馬蹄形に曲がった6つの薄い細胞層である。しかし、そこに投射してくる場所は正確にきまっている。それゆえ、“投射の方向線”の中の細胞柱は、両眼視の視野のうち反対側の視野に関係する各眼の網膜の中の対応する点からの入力を受ける。両眼視の像の融合は、外側膝状体の中では起こらない。それは、網膜膝状体線維が外側膝状体の異なった層に終わっているからである。(術後長期間生存させた例の)視神経の切片を追って調べると、順応性の変性、またはニューロン越えの変性は、各側の外側膝状体の3つの層に起こる。神経線維、または細胞の変性が起こる層は、網膜からの交叉性線維(1,4および6層)と非交叉性線維(2,3および5層)の配列に従って異なる。網膜の小さい傷によって、同側、対側の3つの違った層の中に“投射の方向線”に従って、一列に並んで配列した細胞集団に、ニューロン越えの変性が起こる。反対側の単眼視野(単眼性の半月形の視野)は、網膜の内側半のもっとも内側の部分における受容器要素と関係しており、これからは、2層性の部分に終止する交叉性の神経線維のみが出ている。外側膝状体における“投射の方向線”は、有線野に加えた傷害によって外側膝状体の中に現れる逆行性細胞変性の研究からも、明らかにすることができる。視野の半分は、局在性をもってそれぞれの側の半球の有線野に部位局在的に投射しているので、外側膝状体の中に逆行性細胞変性が現れる帯状の部分は、それぞれの側で、投射の方向線によって境される。外側膝状体では視野の中心を通る水平線が背腹方向に入る斜めの面に対応し、この面により内側部と外側部が分けられる。両眼の網膜の上半部からの神経線維は、外側膝状体の内側半部に投射し、下半分は、本核の外側半部に線維を送る。網膜の黄斑からの投射は、外側膝状体尾側部のうち視野の中心を水平子午線に相当する面の両側にある。幅の広いクサビ形の部分として示される。黄斑に相当する部分は、外側膝状体の全容積のおよそ12%を占める。視野の中心を通る垂直の子午線に一致し、本核の尾側縁に沿って、内側の境界からの外側境界に及んでいる。外側膝状体は、視索の主要な終止場所である。ここから膝状体鳥距路、あるいは視放線を経て、鳥距周囲皮質(17野)に投射があり、また外側膝状体は、この皮質野から皮質膝状体線維を受ける。本核は、視床枕と核間結合をしている。)
- 871_23【Optic tract視索 Tractus opticus】 Portion of the visual pathway between the optic chiasm and lateral geniculate body that is visible on the surface of the basal part of the brain.
→(視索は視交叉と外側膝状体の間の視覚路で左右の視索は視床下部と大脳脚基底部を回って後外方へ走る。これらの線維の多くは外側膝状体の中に終止するが、小部分は下丘腕となって上丘および視蓋前域にまで続く。外側膝状体からは膝状体鳥距路が起こり、これが視覚路の最後の中継路をなす。視索前域は対光反射と関係し、上丘は眼と頭の反射運動より視覚刺激を追跡することと関係している。網膜視床下部線維は、両側性に視床下部の視神経交叉上核に終止する。この網膜からの直接の投射は、機能的には神経内分泌調節と関連している。)
- 871_24【Cerebral peduncle大脳脚[広義の] Pedunculus cerebri】
→(広義の大脳脚は中脳の腹側部で、背側の中脳蓋(四丘体)および中心灰白質背側部を除いた中脳水道水平中央断面より腹側の部分を総称する。さらにこれは背側の中脳被蓋と狭義の大脳脚に分かれる。中脳被蓋には著明な構造物として、動眼神経核群、中脳網様体、赤核、黒質、内側毛帯などが存在する。もともとは全脳と後脳を連結するやや細くなった首状部分である中脳の両半分の部分をさす名称であったが、その後、様々な意味で用いられるようになった。Crus cerebriとよばれる皮質投射線維の大きな束のみをさしたり、これに被蓋を加えたものをさしたりするが、後者の方が好ましい、脚底にある黒質は被蓋とcrus cerebriとを境する構造とみなされている。)