(中枢神経系の神経核)Nuclei systematis nervosi cenralis [A17_2_00_001]
Cranial nerve nuclei(脳神経核)Nuclei nervorum canialium (encephalicorum) のうしんけいかく
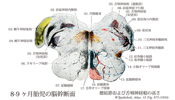
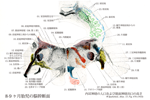
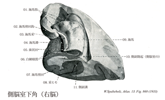
[A17_2_01_001]→脊髄では前角は運動線維の起始領域であり、後角が知覚線維の終末領域であるが、このような脊髄での関係と同様に、脳幹の脳神経核でも遠心性線維を出す細胞のある起始核と、脳幹の外の知覚神経節の偽単極細胞の突起である求心性線維の終わる終止核とが見られる。 体運動性の核は正中線のそばにある:舌下神経核(舌筋へ);外転神経核、滑車神経核、動眼神経核。後者はいずれも眼筋へいく神経の起始核。 それらの外側に接して内臓運動性の核がある。真に内臓運動性で副交感神経に属する核と、かつては内臓運動性であったが、今では変化した鰓弓筋を支配する核とがある。副交感神経核に属するものは、迷走神経背側核(内臓へ行く)、下唾液核(舌下腺へ行く節前線維を出す)、上唾液核(顎下及び舌下腺へ行く節前線維を出す)およびエディンガーウェストファル核(瞳孔括約筋と毛様体筋への節前線維を出す)である。 鰓弓神経の運動核の系列は、下方では副神経(肩の筋を支配)、すんわち頚髄へまで伸びている副神経脊髄根核にはじまる。この核の系列は上方へは疑核すなわち迷走神経と舌咽神経の運動核(咽頭と喉頭の筋を支配する)、および顔面神経核(顔面筋を支配)へと続いていく。顔面神経核はすべての鰓弓神経の運動核と同じく深部にある。この顔面神経線維は背方へ向かい、菱形窩の底(顔面神経丘)で弓形を描いて外転神経核を内側から外側へまわって(顔面神経膝)、その後再び橋の下縁まで下行し、ここで脳幹から出て行く。鰓弓神経の最も上方にある運動核は三叉神経運動核(咀嚼筋へ行く)である。 さらに外側には知覚核が位置し知恵いる:そのうち内側のものは内臓知覚性の孤束核であり、ここには迷走神経と舌咽神経およびその他すべての味覚線維芽終わっている。さらに外側には三叉神経の核領域が広がっており、この領域には三叉神経主知覚核、三叉神経中脳路核、および三叉神経脊髄路核があって、すべての脳神経核のうちで最大の広がりをもっている。この領域には顔面、口および上顎洞からの外受容性感覚のすべての線維芽終わっている。最後に、最外側には前庭神経核と蝸牛神経核のある小野が位置しており、ここには前庭神経(平衡感覚器からくる)および蝸牛神経(聴覚器からくる)が終わっている。
Olfactory glomerulus(嗅糸球体;糸球(嗅球の))Glomerulus olfactorius; Glomerula olfactoria きゅうしきゅうたい;しきゅう(きゅうきゅうの)
[A17_2_01_002]→嗅球内の小弓状領域の1つで、その中では僧帽細胞と房飾細胞の樹状突起が嗅覚受容細胞の軸索とシナプスする。
Nucleus of oculomotor nerve(動眼神経核)Nucleus nervi oculomotorii
[A17_2_01_003]→動眼神経核は第三脳神経、すなわち、動眼神経の起始核であり、動眼神経運動神経細胞の集合である。中脳被蓋の正中線背側部の両側に存在し、中脳中心灰白質の腹方に位置する。動眼神経核の尾側端は滑車神経核の吻側端とほとんど連続しており、動眼神経核の吻側端は中脳の最吻側レベルに達する。動眼神経運動神経細胞はそれぞれの筋支配に対応して局在配列する。すなわち、下直筋支配細胞は核の背側部、内直筋支配細胞は核の腹側部、下斜筋支配細胞は前2者の中間部、上直筋支配細胞は内側部に位置し、それぞれ脳幹の長軸に平行な左右一対の細胞柱を形成する。これらのうち、上直筋支配細胞柱だけは反対側の筋を支配するが、その他は同側の筋を支配する。また、上眼瞼挙筋を支配する細胞柱は正中部に位置し、不対であって、動眼神経核の尾側1/3のレベルにのみ存在する(尾側正中核)。以上の他、副核(Edinger-Westphal核)および正中核(Perlia核)を含めて動眼神経核群とされることがおおい。
Posterior nucleus of accessory oculomotor nuclei; ; Dorsal nucleus of accessory oculomotor nuclei; Dorsal visceral nucleus; Posterior visceral nucleus; Posterior accessory nucleus of optic tract(背側核;背外側核;後核(動眼神経副核の))Nucleus dorsalis accessorii nervi oculomotorii; Nucleus posterior accessorii tracti optici
[A17_2_01_004]→
Anterior medial nucleus visceral nucleus; Ventral medial visceral nucleus(前内側核;腹内側核;前正中核)Nucleus anteromedialis accessorii nervi oculomotorii; Nucleus visceralis anteromedialis; Nucleus visceralis dorsalis
[A17_2_01_005]→左右のEdinger-Westphal核は上方では正中線の背側で合一し、前正中核の内臓性細胞と一続きになる。前正中核の細胞は上部の外側体性細胞柱間の縫線内にある。Edinger-Westphal核および前正中核の両者からは非交叉性の副交感神経節前線維が出て、体性線維と共に走って網様体神経節に至る。内臓性核はこれまで毛様体神経節に至る副交感神経節前線維のみを投射すると考えられてきたが、最近の逆行性軸索輸送法を用いた研究により下位脳幹や脊髄にも投射するすることが明らかにされた。
Central caudal nucleus of oculomotor nerve(正中核;動眼神経正中核)Nucleus caudalis centralis nervi oculomotorii (Perlia) せいちゅうかく;どうがんしんけいせいちゅうかく
[A17_2_01_006]→
Accessory nuclei of oculomotor nerve(動眼神経副核;副起始核(動眼神経の);エディンガー・ウェストファール核)Nuclei accessorii nervi olulomotorii
[A17_2_01_007]→動眼神経副核;副起始核(動眼神経の);エディンガー・ウェストファール核#TA(html)\A14_1_06_303.html#
Nucleus of trochlear nerve; Trochlear nucleus(滑車神経核)Nucleus nervi trochlearis
[A17_2_01_008]→滑車神経核は下丘の高さで、中心灰白質の腹内側にあり、動眼神経核の直接尾方延長部にあたる。これは動眼神経主核と同様に多極性の大細胞からなり、これから出る根線維は中心灰白質の外側部を背側方でやや下方に走り、下丘の下で完全交叉をおこない(滑車神経交叉)、脳を出る。なお滑車神経核の背側で中心灰白質中には背側被蓋核が認められ、また内側縦束の腹側には腹側被蓋核が区別される。
Motor nucleus of facial nerve(三叉神経核)Nucleus nervi facialis
[A17_2_01_009]→三叉神経核は咀嚼筋を支配している運動核と主にその固有感覚に関与する中脳路核、および顔面、口腔内領域と脳膜の一部の一般体性感覚に関与する感覚核群からなる。感覚核群はさらに主感覚核および脊髄路核からなる。運動核は橋の腹外側から脳幹を出て感覚根の内側部に沿って走り、三叉神経節の下部を通って咀嚼筋にいたる。感覚核群のうち主感覚核は橋の高さで運動核の背外側方に位置して存在する。主感覚核より下方につづいて脊髄路核が存在するが、これは延髄の全長にわたり背外側に位置し、下方に頚髄第2節の高さにまで伸び、次第に頚髄後核に移行する。この核は細胞構築的に上方より上位核、中位核および下核に細分される。感覚核群は主感覚核および脊髄路核ともに三叉神経節(半月神経節)の中枢枝の線維を受ける。脊髄路核にいたるこれらの線維は、核の外側方をおおって三叉神経脊髄路を形成している。三叉神経脊髄路内では末梢の三枝からの局在が明確で、腹側より眼枝、上顎枝および下顎枝からの線維がその順に背側方に向けて並ぶ。さらに脊髄路の背内側端には舌咽および迷走神経の中枢枝が加わる。感覚群からは第2次経路としてそれぞれの高さから視床に向かう線維がでる。主感覚核の背側部から出た線維は中心被蓋路に沿って上行枝の視床後内腹側核におわる。主感覚核の腹側部および脊髄路核から視床へ向かう線維が出る。主感覚核の背側部からでた線維は中心被蓋路に沿って上行し同側の視床後内腹側核におわる。主感覚核の腹側部および脊髄路核から視床へ向かう線維はそれぞれの高さで交叉し、対側の内側毛帯と併走して三叉神経毛帯をなして対側の視床後内側腹側核にいたる。上記の視床にいたる線維のほかに、感覚核群からは両側の網様体、対側の上丘および小脳に線維を送る。機能的には主感覚核は主に触圧覚に関与し、脊髄路核ことに下核は温痛核に関与するといわれる。 第7脳神経、すなわち顔面[中間]神経を形成する神経線維のうち、表情筋・広頚筋・アブミ骨筋・茎突舌骨筋などの横紋筋を支配する運動神経線維の起始核であり、橋の最尾側レベルにおいて橋被蓋の腹外側部に位置する。顔面神経核からおこる神経線維は核の背側から出て背内頭側に走り(顔面神経上行根)、第四脳室底の直下で外転神経核の内側部に達してはじめて密な神経束を形成する。ついで、この線維束は外転神経核の頭側レベルで核の背側を外側に向かい顔面神経膝を形成する。ついで、線維束は三叉神経脊髄路核の内側縁に沿うように腹外側に走り(顔面神経下行根)、橋の尾側レベルで脳幹を出る。顔面神経の支配を受ける横紋筋のうち顎二腹筋後腹は副顔面神経核に支配される。副顔面神経核の神経細胞は顔面神経核と三叉神経運動核を結ぶ線上に散在性に存在する。顔面神経に含まれる副交感神経線維の起始核として、上唾液核が記載されている。この核の神経細胞は、橋被蓋網様体の尾側レベルでその背外側部において、三叉神経脊髄路核の内側縁付近に比較的散在性に存在するようである。
Motor nucleus of trigeminal nerve(三叉神経運動核;三叉神経起始核)Nucleus motorius nervi trigemini
[A17_2_01_010]→三叉神経運動核は典型的な大型の神経細胞よりなり、全体は卵円形をなして運動根と主知覚核の内側に位置する。核からの線維は知覚根の進入点より内側で脳幹を出て三叉神経節の下を通り、下顎神経に加わる。運動核は中脳路核からの側副枝を受けて2ニューロン反射弓を形成するが、そのほかにも三叉神経二次線維が交叉性および非交叉性に連絡しており、皮膚、舌および口腔粘膜と咀嚼筋との間に反射弓を形成する。皮質延髄路線維の一部は直接両側性に運動核細胞に終止するが、他の線維は網様体細胞を介して間接的に終止する。
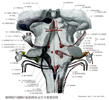
Principal sensory nucleus of trigeminal nerve; Superior sensory nucleus of trigeminal nerve(三叉神経主感覚核;三叉神経主知覚核;三叉神経上知覚核)Nucleus principalis nervi trigemini; Nucleus pontinus nervus trigemini
[A17_2_01_011]→三叉神経主知覚核は上知覚核ともよばれ、外転神経核の上外側にあり、下方は脊髄路核に接し、円い中等大細胞の不規則なぶどう状の集団からなる。主知覚核およびおそらくは脊髄路核の上部は、四肢および体幹に対する後索核と同様に、顔面の識別性触圧覚を中継する物と考えられる。
Mesencephalic nucleus of trigeminal nerve(三叉神経中脳路核;三叉神経中脳核)Nucleus mesencephalicus nervi trigeminalis
[A17_2_01_012]→三叉神経中脳路核は三叉神経運動核の下端より少し下方の高さから中脳上端部に至る。非常に細長く延びた核で、橋の高さでは第四脳室の腹外側核の近くにあり、中脳では中心灰白質の外側縁にある。これは脊髄神経節の細胞に似た、少数の大きい偽単極細胞からなる。その突起は三叉神経中脳路を作りつつ下行し、三叉神経運動核に突起を出したのち運動根に加わり、主として咀嚼筋に分布し、その固有知覚を伝え、咬む力の調節に関与している。また中脳路核上部には外眼筋などからの固有知覚が伝えられるという。中脳路核から高次の中枢への連絡については不明である。
Motor nucleus of facial nerve(三叉神経脊髄路核)Nucleus nervi facialis
[A17_2_01_013]→第7脳神経、すなわち顔面[中間]神経を形成する神経線維のうち、表情筋・広頚筋・アブミ骨筋・茎突舌骨筋などの横紋筋を支配する運動神経線維の起始核であり、橋の最尾側レベルにおいて橋被蓋の腹外側部に位置する。顔面神経核からおこる神経線維は核の背側から出て背内頭側に走り(顔面神経上行根)、第四脳室底の直下で外転神経核の内側部に達してはじめて密な神経束を形成する。ついで、この線維束は外転神経核の頭側レベルで核の背側を外側に向かい顔面神経膝を形成する。ついで、線維束は三叉神経脊髄路核の内側縁に沿うように腹外側に走り(顔面神経下行根)、橋の尾側レベルで脳幹を出る。顔面神経の支配を受ける横紋筋のうち顎二腹筋後腹は副顔面神経核に支配される。副顔面神経核の神経細胞は顔面神経核と三叉神経運動核を結ぶ線上に散在性に存在する。顔面神経に含まれる副交感神経線維の起始核として、上唾液核が記載されている。この核の神経細胞は、橋被蓋網様体の尾側レベルでその背外側部において、三叉神経脊髄路核の内側縁付近に比較的散在性に存在するようである。
Nucleus of abducens nerve; Abducens nucleus (VI)(外転神経核)Nucleus nervi abducentis
[A17_2_01_014]→第6脳神経、すなわち、外転神経の起始核で、眼筋のうち外側直筋を支配する運動神経細胞群である。第四脳室底において橋の正中線背側核の両側に位置しており、顔面神経膝とともに、菱形窩に低い隆まりを形成する。
Nuclei of solitary tract; Solitary nuclei(孤束核)Nuclei tractus solitarii
[A17_2_01_015]→孤束核は延髄背側を矢状方向に走る細い細胞柱で、菱形窩底の下で境界溝のすぐ外側にある。小さい独立核の集まりで一括して言えば脳幹の内臓求心性の核であって迷走神経、舌咽神経、顔面神経からの線維が孤束を経てはいってくる。孤束核は次の数個の部分に分けられる。すなわち、①内側部、迷走神経背側運動核の背外側にある、②背内側、背外側および腹外側亜核、孤束を取り巻く、および③小細胞性亜核、最後野の腹側にある。内側部の細胞は迷走神経背側運動核を吻側にわずかに越え、また、第四脳室より下方へ広がり、反対側の同じ細胞性と合一して迷走神経交連核をつくる。外側の諸核は大細胞性の細胞柱をなし、孤束を一部あるいは完全に取り巻いている。この外側部は孤束のほぼ全長にわたって平行に存在する。吻側では橋下部まで広がり、一方尾側ではその細胞数が減少し、網様体との区別が難しくなる。孤束核の膨大した吻側部(すなわち外側部)は主として特殊内臓求心性(味覚)線維を顔面神経(中間神経)と一部の顔面神経と舌咽神経からくる一般内臓求心性線維を受ける。孤束核の内側部の細胞は多数のニューロペプチドを含有している。すなわち、エンケファリン、ソマトスタチン、サブスタンスP、およびコレシストキニンなどである。同部位にはサブスタンスP含有線維も豊富に存在する。孤束核から起こる二次路系は次の部位に同側性に投射する。すなわち、①疑核とその周辺の網様体、②橋上部の傍腕核、および③味覚に関与する視床の核、すなわち後内側腹側核(小細胞部)である。孤束核から起こるその他の二次線には舌下神経核と唾液核に投射し、舌の運動と唾液の分泌反射を仲介する。孤束核からの迷走神経背側運動核、横隔神経核(第三(C3)・第四(C4)・第五(C5)頚髄の高さ)、および胸髄の前角細胞への投射はせきと嘔吐反射に関与する。孤束核は生理学的に同定された延髄の呼吸“中枢”と同一の広がりをもち、この中枢は疑核とその周辺の網様体を包含する。延髄“呼吸中枢”の細胞は迷走神経のインパルス、および直接の化学的環境変化(CO2蓄積)によって賦活される。別個の昇圧帯と減圧帯からなる延髄の血管運動“中枢”は十分明らかにされていない。最近、心臓血管系の調節に関する神経回路網が重視されてきた。ノルアドレナリン作働性ニューロンの1集団(グループA5と呼ばれる)が橋の下部で上オリーブ核と顔面神経の根線維の間にあり、軸索を孤束核、疑核、迷走神経背側運動核および胸髄の交感神経節前線維のニューロンに投射しており、上記神経回路網の一部と思われる。
Superior salivary nucleus; Superior salivatory nucleus(上唾液核;中間神経分泌核)Nucleus salivarius superior; Nucleus secretorius nervi intermedii
[A17_2_01_016]→上唾液核は舌下腺、顎下腺、口蓋腺などの唾液腺や涙腺の分泌を行う中間神経副交感性線維の起始核である。起始細胞は、延髄上部の毛様体外側部で肺内層から腹外側方向に配列されている。尾側端は顔面神経核の高さで孤束の腹内側に位置し、吻側端は上オリーブ核の高さで前庭神経核の腹側から三叉神経脊髄路核の内側に位置している。
Nuclei of solitary tract; Solitary nuclei(孤束核)Nuclei tractus solitarii
[A17_2_01_017]→孤束核は延髄背側を矢状方向に走る細い細胞柱で、菱形窩底の下で境界溝のすぐ外側にある。小さい独立核の集まりで一括して言えば脳幹の内臓求心性の核であって迷走神経、舌咽神経、顔面神経からの線維が孤束を経てはいってくる。孤束核は次の数個の部分に分けられる。すなわち、①内側部、迷走神経背側運動核の背外側にある、②背内側、背外側および腹外側亜核、孤束を取り巻く、および③小細胞性亜核、最後野の腹側にある。内側部の細胞は迷走神経背側運動核を吻側にわずかに越え、また、第四脳室より下方へ広がり、反対側の同じ細胞性と合一して迷走神経交連核をつくる。外側の諸核は大細胞性の細胞柱をなし、孤束を一部あるいは完全に取り巻いている。この外側部は孤束のほぼ全長にわたって平行に存在する。吻側では橋下部まで広がり、一方尾側ではその細胞数が減少し、網様体との区別が難しくなる。孤束核の膨大した吻側部(すなわち外側部)は主として特殊内臓求心性(味覚)線維を顔面神経(中間神経)と一部の顔面神経と舌咽神経からくる一般内臓求心性線維を受ける。孤束核の内側部の細胞は多数のニューロペプチドを含有している。すなわち、エンケファリン、ソマトスタチン、サブスタンスP、およびコレシストキニンなどである。同部位にはサブスタンスP含有線維も豊富に存在する。孤束核から起こる二次路系は次の部位に同側性に投射する。すなわち、①疑核とその周辺の網様体、②橋上部の傍腕核、および③味覚に関与する視床の核、すなわち後内側腹側核(小細胞部)である。孤束核から起こるその他の二次線には舌下神経核と唾液核に投射し、舌の運動と唾液の分泌反射を仲介する。孤束核からの迷走神経背側運動核、横隔神経核(第三(C3)・第四(C4)・第五(C5)頚髄の高さ)、および胸髄の前角細胞への投射はせきと嘔吐反射に関与する。孤束核は生理学的に同定された延髄の呼吸“中枢”と同一の広がりをもち、この中枢は疑核とその周辺の網様体を包含する。延髄“呼吸中枢”の細胞は迷走神経のインパルス、および直接の化学的環境変化(CO2蓄積)によって賦活される。別個の昇圧帯と減圧帯からなる延髄の血管運動“中枢”は十分明らかにされていない。最近、心臓血管系の調節に関する神経回路網が重視されてきた。ノルアドレナリン作働性ニューロンの1集団(グループA5と呼ばれる)が橋の下部で上オリーブ核と顔面神経の根線維の間にあり、軸索を孤束核、疑核、迷走神経背側運動核および胸髄の交感神経節前線維のニューロンに投射しており、上記神経回路網の一部と思われる。
Nucleus of vestibulocochlear nerve(内耳神経核)Nuclei nervi vestibulocochlearis ないじしんけいかく
[A17_2_01_018]→内耳神経核は菱形窩の中央部の外側部から下方にわたって存在し、前庭神経核と蝸牛神経核とに分けられる。(解剖学講義)
Anterior cochlear nucleus; Ventral cochlear nucleus(蝸牛神経腹側核)Nucleus cochlearis anterior
[A17_2_01_019]→蝸牛神経腹側核は発育がよく、下小脳脚の腹外側で、蝸牛神経の根の外側にある。蝸牛神経腹側核はその部位と細胞構築によって、前腹側核と後腹側核の2亜核に分けられる。蝸牛神経腹側前核anteroventral cochlear nucleusは吻側で卵円形細胞が密につまっており、また蝸牛神経腹側後核posteroventral cochlear nucleusは蝸牛神経の脳内侵入部近くにあって種々の形の神経細胞よりなるが、そのうちの主な物は多極性である。これらの亜核には音階的配列があり、音階に応じて連続的に対応している。蝸牛神経の線維は脳幹内に入ったのち、規則正しく順番に分岐して蝸牛神経背側核および腹側核の両方に音快的配列をもって終止する。蝸牛神経核内で線維が次々と規則正しく分岐と分布を行うので全ての亜核内で重なりあった周波数配列が認められる。蝸牛神経核では高い周波数に反応する細胞が背側に、低い周波数に反応する細胞が腹側に位置するが、これは蝸牛とは反対の配列である。
Posterior cochlear nucleus; Dorsal cochlear nucleus(蝸牛神経背側核)Nucleus cochlearis posterior
[A17_2_01_020]→蝸牛神経背側核は腹側核よりやや下方の高さで下小脳脚の背外側に接し、菱形窩の聴結節にある。この核は多くの動物では層構造を示すが、ヒトでも多少その傾向が認められる。
Medial vestibular nucleus in pontile tegmentum(前庭神経内側核;前庭神経三角核(橋被蓋の))Nucleus vestibularis medialis; Nucleus terminalis triangularis nervus vestibuli
[A17_2_01_021]→前庭神経核群のうち、前庭神経下核は延髄において副楔状束核の内側から上方へ前庭神経が脳幹に入るまでの高さにあり、主として小型および中型の細胞よりなる。ただし、その最も吻側部は前庭神経外側核に似た大型細胞から成る。神経線維染色標本で縦走する線維束の認められるのがこの核の特徴である。
Lateral vestibular nucleus(前庭神経外側核)Nucleus vestibularis lateralis
[A17_2_01_022]→前庭神経外側核はダイテルス核ともよばれる。前庭神経核は4つの小核からなるが、その内の外側核をいう。前庭神経外側核は前庭神経が脳幹内に入る高さにあり、巨大型細胞よりなるが、細胞の数および形には部位的差異が認められる。前庭神経の根線維はこの核の腹側部を通る。ドイツの解剖学者Otto Friedrich Karl Deiters (1834-1863)によって記載された。
Superior vestibular nucleus(前庭神経上核;前庭神経背側核)Nucleus vestibularis superior; Nucleus terminalis dorsalis nervus vestibuli
[A17_2_01_023]→ベヒテレフ核とも呼ばれる。前庭神経上核は前庭神経外側核の上方背側にあり、その背側を上小脳脚の線維が通る。核の中央には大型細胞があり、その周囲をそれより小形の細胞が取り囲んでいる。Bechterew, Vladimir Michaliorich (Bekhterev)(1857-1927)ロシアの神経学者。1893年からペテルスブルグ大学の教授。1918年に同市の脳精神研究所の初代所長となる。ロシアの神経学の先駆者、聴神経のベヒテレフ核、大脳皮質のベヒテレフ層、ベヒテレフ病(強直性脊椎関節炎)を記述(1892年)
Inferior vestibular nucleus(前庭神経下核;前庭神経下行路核)Nucleus vestibularis inferior
[A17_2_01_024]→前庭神経核群のうち、前庭神経下核は延髄において副楔状束核の内側から上方へ前庭神経が脳幹に入るまでの高さにあり、主として小型および中型の細胞溶離成る。ただし、その最も吻側部は前庭神経外側核に似た大型細胞から成る。神経線維染色標本で縦走する線維束の認められるのがこの核の特徴である。
Nucleus of glossopharyngeal nerve(舌咽神経核)Nucleus nervi glossopharyngei
[A17_2_01_025]→舌咽神経核 舌咽神経(第9脳神経)の起始核、終止核のことで以下の4つの核からなる。①下唾液核:迷走神経背側核のすぐ吻側にある一般内臓性遠心性線維の起始核で、その神経細胞の軸索は、副交感神経節前線維として耳神経節で節後線維に中継されて耳下腺の分泌に関与する。②疑核:吻側部の神経細胞の軸索は特殊内臓性遠心性線維として茎突咽頭筋と咽頭収縮筋の上部を支配する。③孤束核:吻側部には舌の後1/3の味覚を伝える特殊内臓性求心性線維が終止している。④舌咽神経背側核:迷走神経背側核の吻外側孤束核の内側に位置する神経核で、一般内臓性求心性線維が終止する。(医学書院医学大辞典:大竹一嘉)
Nuclei of solitary tract; Solitary nuclei(孤束核)Nuclei tractus solitarii
[A17_2_01_026]→孤束核は延髄背側を矢状方向に走る細い細胞柱で、菱形窩底の下で境界溝のすぐ外側にある。小さい独立核の集まりで一括して言えば脳幹の内臓求心性の核であって迷走神経、舌咽神経、顔面神経からの線維が孤束を経てはいってくる。孤束核は次の数個の部分に分けられる。すなわち、①内側部、迷走神経背側運動核の背外側にある、②背内側、背外側および腹外側亜核、孤束を取り巻く、および③小細胞性亜核、最後野の腹側にある。内側部の細胞は迷走神経背側運動核を吻側にわずかに越え、また、第四脳室より下方へ広がり、反対側の同じ細胞性と合一して迷走神経交連核をつくる。外側の諸核は大細胞性の細胞柱をなし、孤束を一部あるいは完全に取り巻いている。この外側部は孤束のほぼ全長にわたって平行に存在する。吻側では橋下部まで広がり、一方尾側ではその細胞数が減少し、網様体との区別が難しくなる。孤束核の膨大した吻側部(すなわち外側部)は主として特殊内臓求心性(味覚)線維を顔面神経(中間神経)と一部の顔面神経と舌咽神経からくる一般内臓求心性線維を受ける。孤束核の内側部の細胞は多数のニューロペプチドを含有している。すなわち、エンケファリン、ソマトスタチン、サブスタンスP、およびコレシストキニンなどである。同部位にはサブスタンスP含有線維も豊富に存在する。孤束核から起こる二次路系は次の部位に同側性に投射する。すなわち、①疑核とその周辺の網様体、②橋上部の傍腕核、および③味覚に関与する視床の核、すなわち後内側腹側核(小細胞部)である。孤束核から起こるその他の二次線には舌下神経核と唾液核に投射し、舌の運動と唾液の分泌反射を仲介する。孤束核からの迷走神経背側運動核、横隔神経核(第三(C3)・第四(C4)・第五(C5)頚髄の高さ)、および胸髄の前角細胞への投射はせきと嘔吐反射に関与する。孤束核は生理学的に同定された延髄の呼吸“中枢”と同一の広がりをもち、この中枢は疑核とその周辺の網様体を包含する。延髄“呼吸中枢”の細胞は迷走神経のインパルス、および直接の化学的環境変化(CO2蓄積)によって賦活される。別個の昇圧帯と減圧帯からなる延髄の血管運動“中枢”は十分明らかにされていない。最近、心臓血管系の調節に関する神経回路網が重視されてきた。ノルアドレナリン作働性ニューロンの1集団(グループA5と呼ばれる)が橋の下部で上オリーブ核と顔面神経の根線維の間にあり、軸索を孤束核、疑核、迷走神経背側運動核および胸髄の交感神経節前線維のニューロンに投射しており、上記神経回路網の一部と思われる。
Nucleus ambiguus(疑核;舌咽迷走神経腹側核)Nucleus ambiguus; Nucleus originis ventralis nervi glossopharyngici et nervi vagi
[A17_2_01_027]→疑核はオリーブの背側にある核。舌咽神経(IX)、迷走神経(X)および副神経(XI)延髄根の起始核。疑核は三叉神経脊髄路核と下オリーブ核群のほぼ中間で、網様体内にある細胞柱である。この核は毛帯交叉の高さより下オリーブ核群の吻側1/3部の高さまで広がり多極性のコリン作働性の下位運動ニューロンからなる。この核より出た線維は背側に弧を描き、迷走神経背側運動核からの遠心線維と一緒になり下オリーブ核群の背側の延髄外側面から外にでる。疑核の尾方部は副神経の延髄根をだし、吻側部は舌咽神経の特殊内臓遠心性線維をだす(茎突咽頭筋を支配する)。迷走神経の特殊内臓遠心性線維(迷走神経に結合する副神経延髄根からの線維を含めて)は咽頭と喉頭の筋を支配する。疑核の細胞はNissl染色標本では同定が困難であり、この細胞は線維を出し、副神経脊髄根の一部を構成する。疑核の細胞はコリン作働性であるので、コリンアセチルトランスフェラーゼ(ChAT)により免疫細胞化学反応を行えば、容易に見ることができる。舌下神経核と迷走神経背側運動核の細胞もChAT免疫反応陽性である。
Nuclei of solitary tract; Solitary nuclei(孤束核)Nuclei tractus solitarii
[A17_2_01_028]→孤束核は延髄背側を矢状方向に走る細い細胞柱で、菱形窩底の下で境界溝のすぐ外側にある。小さい独立核の集まりで一括して言えば脳幹の内臓求心性の核であって迷走神経、舌咽神経、顔面神経からの線維が孤束を経てはいってくる。孤束核は次の数個の部分に分けられる。すなわち、①内側部、迷走神経背側運動核の背外側にある、②背内側、背外側および腹外側亜核、孤束を取り巻く、および③小細胞性亜核、最後野の腹側にある。内側部の細胞は迷走神経背側運動核を吻側にわずかに越え、また、第四脳室より下方へ広がり、反対側の同じ細胞性と合一して迷走神経交連核をつくる。外側の諸核は大細胞性の細胞柱をなし、孤束を一部あるいは完全に取り巻いている。この外側部は孤束のほぼ全長にわたって平行に存在する。吻側では橋下部まで広がり、一方尾側ではその細胞数が減少し、網様体との区別が難しくなる。孤束核の膨大した吻側部(すなわち外側部)は主として特殊内臓求心性(味覚)線維を顔面神経(中間神経)と一部の顔面神経と舌咽神経からくる一般内臓求心性線維を受ける。孤束核の内側部の細胞は多数のニューロペプチドを含有している。すなわち、エンケファリン、ソマトスタチン、サブスタンスP、およびコレシストキニンなどである。同部位にはサブスタンスP含有線維も豊富に存在する。孤束核から起こる二次路系は次の部位に同側性に投射する。すなわち、①疑核とその周辺の網様体、②橋上部の傍腕核、および③味覚に関与する視床の核、すなわち後内側腹側核(小細胞部)である。孤束核から起こるその他の二次線には舌下神経核と唾液核に投射し、舌の運動と唾液の分泌反射を仲介する。孤束核からの迷走神経背側運動核、横隔神経核(第三(C3)・第四(C4)・第五(C5)頚髄の高さ)、および胸髄の前角細胞への投射はせきと嘔吐反射に関与する。孤束核は生理学的に同定された延髄の呼吸“中枢”と同一の広がりをもち、この中枢は疑核とその周辺の網様体を包含する。延髄“呼吸中枢”の細胞は迷走神経のインパルス、および直接の化学的環境変化(CO2蓄積)によって賦活される。別個の昇圧帯と減圧帯からなる延髄の血管運動“中枢”は十分明らかにされていない。最近、心臓血管系の調節に関する神経回路網が重視されてきた。ノルアドレナリン作働性ニューロンの1集団(グループA5と呼ばれる)が橋の下部で上オリーブ核と顔面神経の根線維の間にあり、軸索を孤束核、疑核、迷走神経背側運動核および胸髄の交感神経節前線維のニューロンに投射しており、上記神経回路網の一部と思われる。
Dorsal nucleus of glossopharyngeal nerve(舌咽神経背側核)Nucleus dorsalis nervi glossopharyngei ぜついんしんけいはいそくかく
[A17_2_01_029]→迷走神経背側核はさらに内側核と外側核とに分けられる。内側核(背側運動核dorsal motor nucleus)は一般内臓遠心性である副交感性線維を出す。外側核は迷走神経支配の領域から一般内臓求心線維が終止する。この核はまた孤束核に含められ、孤束核の内側核として取り扱われることが多い。迷走神経背側運動核は舌下神経核の背外側で第四脳室底に位置する。比較的小さい紡錘形の細胞からなるこの細胞柱は、吻側および尾側共にわずかに舌下神経核を越えて広がっている。この神経核の一部の大きい細胞は粗大なNissl小体と散在性のメラニン色素を含む。迷走神経背側運動核の細胞は副交感神経節前線維(GVE)をだし、軸索は軸索は三叉神経脊髄路およびその核を貫いて延髄外側面より外にでる。迷走神経背側運動核迷走神経性の分泌運動中枢である。迷走神経背側運動核の細胞はコリンアセチルトランスフェラーゼ(ChAT)に強く免疫陽性を示す。この核を破壊すると、インスリンによって誘発される胃液分泌が大きく減弱する。
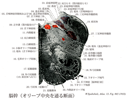
Nucleus of vagus nerve(迷走神経核(灰白翼))Nucleus nervi vagi (alae cinereae); Nucleus alae cinereae
[A17_2_01_029_1]→第四脳室底の迷走神経三角の部分に存在する、遠心性線維の起始核と求心性線維の終止核からなり次の三つの核が区別される。(1)迷走神経背側核:これはさらに内側核と外側核とにわけられる。内側核(背側運動核)は一般内臓遠心性である副交感性線維を出す。外側核は迷走神経支配の領域からの一般内臓求心性線維が終止する。この核はまた孤束核に含められ、孤束核の内側核として扱われることが多い。(2)孤束核:孤束の外側にある大型細胞の集団で喉頭蓋領域からの特殊内臓求心性線維である味覚線維が終わる。(3)疑核:延髄網様体でオリーブ核と三叉神経脊髄路核との間に位置する運動性細胞群である。特殊内臓遠心性線維の起始核で、茎突咽頭筋を除く咽頭筋や軟口蓋の筋などの横紋筋を支配する。疑核の吻側部は茎突咽頭筋支配の舌咽神経の特殊内臓遠心線維を、尾側部は副神経延髄根を出す。なお背側核や疑核の周辺には心臓運動抑制線維を出す細胞がある。耳介、外耳道、鼓膜などからの迷走神経の一般体性求心性線維は三叉神経脊髄路核に終わる。(解剖学辞典:松下松雄)
Nucleus of vagus nerve(迷走神経核)Nucleus nervi vagi
[A17_2_01_030]→第四脳室底の迷走神経三角の部分に存在する、遠心性線維の起始核と求心性線維の終止核からなり次の三つの核が区別される。(1)迷走神経背側核:これはさらに内側核と外側核とにわけられる。内側核(背側運動核)は一般内臓遠心性である副交感性線維を出す。外側核は迷走神経支配の領域からの一般内臓求心性線維が終止する。この核はまた孤束核に含められ、孤束核の内側核として扱われることが多い。(2)孤束核:孤束の外側にある大型細胞の集団で喉頭蓋領域からの特殊内臓求心性線維である味覚線維が終わる。(3)疑核:延髄網様体でオリーブ核と三叉神経脊髄路核との間に位置する運動性細胞群である。特殊内臓遠心性線維の起始核で、茎突咽頭筋を除く咽頭筋や軟口蓋の筋などの横紋筋を支配する。疑核の吻側部は茎突咽頭筋支配の舌咽神経の特殊内臓遠心線維を、尾側部は副神経延髄根を出す。なお背側核や疑核の周辺には心臓運動抑制線維を出す細胞がある。耳介、外耳道、鼓膜などからの迷走神経の一般体性求心性線維は三叉神経脊髄路核に終わる。(解剖学辞典:松下松雄)
Nucleus ambiguus(疑核)Nucleus ambiguus
[A17_2_01_031]→疑核はオリーブの背側にある核。舌咽神経(IX)、迷走神経(X)および副神経(XI)延髄根の起始核。疑核は三叉神経脊髄路核と下オリーブ核群のほぼ中間で、網様体内にある細胞柱である。この核は毛帯交叉の高さより下オリーブ核群の吻側1/3部の高さまで広がり多極性のコリン作働性の下位運動ニューロンからなる。この核より出た線維は背側に弧を描き、迷走神経背側運動核からの遠心線維と一緒になり下オリーブ核群の背側の延髄外側面から外にでる。疑核の尾方部は副神経の延髄根をだし、吻側部は舌咽神経の特殊内臓遠心性線維をだす(茎突咽頭筋を支配する)。迷走神経の特殊内臓遠心性線維(迷走神経に結合する副神経延髄根からの線維を含めて)は咽頭と喉頭の筋を支配する。疑核の細胞はNissl染色標本では同定が困難であり、この細胞は線維を出し、副神経脊髄根の一部を構成する。疑核の細胞はコリン作働性であるので、コリンアセチルトランスフェラーゼ(ChAT)により免疫細胞化学反応を行えば、容易に見ることができる。舌下神経核と迷走神経背側運動核の細胞もChAT免疫反応陽性である。
Posterior nucleus of vagus nerve; Dorsal nucleus of vagas nerve(迷走神経背側核)Nucleus posterior nervi vagi; Nucleus dorsalis nervi vagi
[A17_2_01_032]→迷走神経背側核はさらに内側核と外側核とに分けられる。内側核(背側運動核dorsal motor nucleus)は一般内臓遠心性である副交感性線維を出す。外側核は迷走神経支配の領域から一般内臓求心線維が終止する。この核はまた孤束核に含められ、孤束核の内側核として取り扱われることが多い。迷走神経背側運動核は舌下神経核の背外側で第四脳室底に位置する。比較的小さい紡錘形の細胞からなるこの細胞柱は、吻側および尾側共にわずかに舌下神経核を越えて広がっている。この神経核の一部の大きい細胞は粗大なNissl小体と散在性のメラニン色素を含む。迷走神経背側運動核の細胞は副交感神経節前線維(GVE)をだし、軸索は軸索は三叉神経脊髄路およびその核を貫いて延髄外側面より外にでる。迷走神経背側運動核迷走神経性の分泌運動中枢である。迷走神経背側運動核の細胞はコリンアセチルトランスフェラーゼ(ChAT)に強く免疫陽性を示す。この核を破壊すると、インスリンによって誘発される胃液分泌が大きく減弱する。
Nucleus ambiguus(疑核;舌咽迷走神経腹側核)Nucleus ambiguus; Nucleus originis ventralis nervi glossopharyngici et nervi vagi
[A17_2_01_033]→疑核はオリーブの背側にある核。舌咽神経(IX)、迷走神経(X)および副神経(XI)延髄根の起始核。疑核は三叉神経脊髄路核と下オリーブ核群のほぼ中間で、網様体内にある細胞柱である。この核は毛帯交叉の高さより下オリーブ核群の吻側1/3部の高さまで広がり多極性のコリン作働性の下位運動ニューロンからなる。この核より出た線維は背側に弧を描き、迷走神経背側運動核からの遠心線維と一緒になり下オリーブ核群の背側の延髄外側面から外にでる。疑核の尾方部は副神経の延髄根をだし、吻側部は舌咽神経の特殊内臓遠心性線維をだす(茎突咽頭筋を支配する)。迷走神経の特殊内臓遠心性線維(迷走神経に結合する副神経延髄根からの線維を含めて)は咽頭と喉頭の筋を支配する。疑核の細胞はNissl染色標本では同定が困難であり、この細胞は線維を出し、副神経脊髄根の一部を構成する。疑核の細胞はコリン作働性であるので、コリンアセチルトランスフェラーゼ(ChAT)により免疫細胞化学反応を行えば、容易に見ることができる。舌下神経核と迷走神経背側運動核の細胞もChAT免疫反応陽性である。
Nuclei of solitary tract; Solitary nuclei(孤束核)Nuclei tractus solitarii
[A17_2_01_034]→孤束核は延髄背側を矢状方向に走る細い細胞柱で、菱形窩底の下で境界溝のすぐ外側にある。小さい独立核の集まりで一括して言えば脳幹の内臓求心性の核であって迷走神経、舌咽神経、顔面神経からの線維が孤束を経てはいってくる。孤束核は次の数個の部分に分けられる。すなわち、①内側部、迷走神経背側運動核の背外側にある、②背内側、背外側および腹外側亜核、孤束を取り巻く、および③小細胞性亜核、最後野の腹側にある。内側部の細胞は迷走神経背側運動核を吻側にわずかに越え、また、第四脳室より下方へ広がり、反対側の同じ細胞性と合一して迷走神経交連核をつくる。外側の諸核は大細胞性の細胞柱をなし、孤束を一部あるいは完全に取り巻いている。この外側部は孤束のほぼ全長にわたって平行に存在する。吻側では橋下部まで広がり、一方尾側ではその細胞数が減少し、網様体との区別が難しくなる。孤束核の膨大した吻側部(すなわち外側部)は主として特殊内臓求心性(味覚)線維を顔面神経(中間神経)と一部の顔面神経と舌咽神経からくる一般内臓求心性線維を受ける。孤束核の内側部の細胞は多数のニューロペプチドを含有している。すなわち、エンケファリン、ソマトスタチン、サブスタンスP、およびコレシストキニンなどである。同部位にはサブスタンスP含有線維も豊富に存在する。孤束核から起こる二次路系は次の部位に同側性に投射する。すなわち、①疑核とその周辺の網様体、②橋上部の傍腕核、および③味覚に関与する視床の核、すなわち後内側腹側核(小細胞部)である。孤束核から起こるその他の二次線には舌下神経核と唾液核に投射し、舌の運動と唾液の分泌反射を仲介する。孤束核からの迷走神経背側運動核、横隔神経核(第三(C3)・第四(C4)・第五(C5)頚髄の高さ)、および胸髄の前角細胞への投射はせきと嘔吐反射に関与する。孤束核は生理学的に同定された延髄の呼吸“中枢”と同一の広がりをもち、この中枢は疑核とその周辺の網様体を包含する。延髄“呼吸中枢”の細胞は迷走神経のインパルス、および直接の化学的環境変化(CO2蓄積)によって賦活される。別個の昇圧帯と減圧帯からなる延髄の血管運動“中枢”は十分明らかにされていない。最近、心臓血管系の調節に関する神経回路網が重視されてきた。ノルアドレナリン作働性ニューロンの1集団(グループA5と呼ばれる)が橋の下部で上オリーブ核と顔面神経の根線維の間にあり、軸索を孤束核、疑核、迷走神経背側運動核および胸髄の交感神経節前線維のニューロンに投射しており、上記神経回路網の一部と思われる。
Nucleus of accessory nerve; Accessory nucleus (XI)(副神経核)Nucleus nervi accessorii
[A17_2_01_035]→副神経核は脊髄の上方6区域(C1-C6)の前角の中央部と外側部を縦に連ねた運動性細胞柱で、ここから副神経がでる。副神経は延髄根(内枝)と脊髄根(外枝)からなる。延髄根は疑核の下端部から起こり迷走神経に合して喉頭(固有)筋や下咽頭収縮筋を支配する。脊髄根は延髄下部から頚髄上部(C5-C6)にかけて存在する副神経脊髄核からおこり僧帽筋と胸鎖乳突筋を支配する。脊髄核は延髄下部では前索の内側部の近くにあるが下方にいくにしたがい外方に移動し、前角外側部に位置するようになる。根は背外方に向かい、側索の背側部を貫いて脊髄を出る。
Nucleus ambiguus(疑核;舌咽迷走神経腹側核)Nucleus ambiguus; Nucleus originis ventralis nervi glossopharyngici et nervi vagi
[A17_2_01_036]→疑核はオリーブの背側にある核。舌咽神経(IX)、迷走神経(X)および副神経(XI)延髄根の起始核。疑核は三叉神経脊髄路核と下オリーブ核群のほぼ中間で、網様体内にある細胞柱である。この核は毛帯交叉の高さより下オリーブ核群の吻側1/3部の高さまで広がり多極性のコリン作働性の下位運動ニューロンからなる。この核より出た線維は背側に弧を描き、迷走神経背側運動核からの遠心線維と一緒になり下オリーブ核群の背側の延髄外側面から外にでる。疑核の尾方部は副神経の延髄根をだし、吻側部は舌咽神経の特殊内臓遠心性線維をだす(茎突咽頭筋を支配する)。迷走神経の特殊内臓遠心性線維(迷走神経に結合する副神経延髄根からの線維を含めて)は咽頭と喉頭の筋を支配する。疑核の細胞はNissl染色標本では同定が困難であり、この細胞は線維を出し、副神経脊髄根の一部を構成する。疑核の細胞はコリン作働性であるので、コリンアセチルトランスフェラーゼ(ChAT)により免疫細胞化学反応を行えば、容易に見ることができる。舌下神経核と迷走神経背側運動核の細胞もChAT免疫反応陽性である。
Spinal nucleus of accessory nerve(副神経脊髄核)Nucleus spinalis nervus accessorii ふくしんけいせきずいかく [A17_2_01_037]→
Nucleus of hypoglossal nerve; Hypoglossal nucleus; Nucleus of CN XII(舌下神経核)Nucleus nervi hypoglossi ぜっかしんけいかく
[A17_2_01_038]→舌下神経核は第12脳神経、すなわち舌下神経の起始核である。脊髄前角の運動神経細胞群の頭側延長部として、延髄において第四脳室底の直下で正中線背側部の両側に存在し、オリーブ核下端部より内耳神経核のレベルにわたる細胞柱(約2cm)を形成する。内舌筋および5個のうち4個の外舌筋を支配している運動神経核。
Diencephalon(間脳)Diencephalon
[A17_2_02_001]→間脳は「間」を意味するギリシャ語の接頭詞diaと、「脳」を意味するencephalonを結合したもの。間脳は中脳の前方で第三脳室を取り囲んだ領域をいう。背側方は側脳室におおわれ、背外側は分界条によって尾状核と境され、外側を内方によって取り囲まれている。前方は室間孔まで伸び、後方は後交連と乳頭体の後方を結ぶ線で中脳被蓋に移行する。間脳はさらに背側視床、視床下部、腹側視床および視床上部に分かれる。背側視床はこれらのうちもっとも大きな部位を占め左右を視床間橋(中間質)によって結ばれる。背側視床と視床上部とを視床脳とよぶことがある。 間脳の発生development of the diencephalon:間脳は間脳胞から発生する。胎生第4週のおわりころにおける脳の原基は、前脳胞・中脳胞・菱脳胞の3脳胞が確立し、前脳胞ではその外側壁の前腹側部から外方に向かって大きい眼胞が膨出している。第5週に入ると(第14段階)、この眼胞の出発部の前背側にあたる部分の外側壁が前外方、ついで背外方に向かって膨出し、左右1対の半球胞を形成する。左右の半球胞を連ねる前脳胞の前背側端部を終脳正中部という。終脳正中部と半球胞とが終脳を形成し、これまでの前脳胞の大部分を占める領域は、これ以降、間脳胞と呼ばれる。 間脳胞ははじめ頭尾(前後)方向に長い管であり、その内腔は間脳室(後に第三脳室)とよばれる。陥凹胞においても、中脳以下の神経管の各部におけると同じく、実質的な神経細胞の形成は左右の外側壁においてのみおこり、背側壁(蓋板)と腹側壁(底板)においては著明な肥厚は見られない。蓋板は単層立方上皮となり、外側から間葉組織によって裏打ちされて、第三脳室脈絡組織となる。 肥厚・増大していく間脳の外側壁の内面には一過性に前後に走る3本の浅い溝(背側から腹側へ1.間脳背側溝、2.間脳中間溝、3.間脳腹側溝)がみとめられ、これによって外側壁は1.視床上部、2.背側視床、3.腹側視床、4.視床下部の4部に区画される。その後の発育において背側視床と腹側視床がとくに高度に発育し、両者が合一して強大な視床を形成する。胚芽層から生じた多数の神経[芽]細胞は外套層および縁帯の各所に集合して前核・内側核・中心核・腹側核・外側核・視床枕核などの視床核を形成する。 視床上部はヒトでははなはだ退化的で、これに属する構造物としては、間脳の後端部における松果体、手綱および手綱三角のみである。松果体は胎生第7週において間脳の蓋板の後端正中部から背後方に向かって生ずる1個の中空の膨出として発生する。 視床下部は間脳の外側壁および腹側壁を埋め、およそ8個に大別される神経核を形成する。 間脳胞の内腔である間脳室は、はじめは円形に近い横断面を示すが、外側壁の発育につれて上下方向に広く左右方向に狭い空間となり、ついには正中矢状断面に一致した左右の幅の非常に狭い裂隙状の腔となる。この間脳室とその前端につづくごく狭い終脳正中部の内腔を合わせて第三脳室という。
Corpus striatum(線条体)Corpus striatum
[A17_2_03_001]→線条体は尾状核とレンズ核を意味する。レンズ核はさらに被殻と淡蒼球に区別される。このうち尾状核と被殻は終脳胞の腹外側に出現する神経節丘より同一の細胞群として発生し、その後、のちに発達してくる内包によって二つの部分に隔てられたものである。尾状核と被殻とは内包に横切って走る栓状の灰白質によって互いに連なり、特に前下方では両者は構造的にも同じ細胞構築をもっている。線条体という名称は内包を横切って尾状核と被殻を結んでいる灰白質によってできる縞目と、さらに、尾状核や被殻の中を走る有髄線維の小束によってできる縞目とに基づくものである。したがって、尾状核と被殻とをまとめて線条体Striatumとよび、淡蒼球をPallidumとよんで対比することが多い。解剖学用では、慣用されてきたStriatumという語とは異なる意味内容をもつ語として、Corpus striatumが採用されているが、日本名ではどちらも「線条体」である点は注意を要する。一方、淡蒼球の発生や細胞弧対句はStriatumとは異なる。淡蒼球は有髄神経線維に富むため黄灰白色を呈し、赤みを帯びた暗灰色のStriatumとは肉眼的にも明らかに識別できる。系統発生的視点に立って、尾状核と被殻を新線条体(Neostriatum)、淡蒼球を古線条体(Paleostriatum)、扁桃体を原線条体(Archistriatum)とよぶことがある。
Thalamus(視床)Thalamus
[A17_2_03_002]→視床は、間脳の大きいほうの背側部分を形成する灰白質。背側間脳溝と視床下溝の間の領域であるが、発生の間に大きく発育して、間脳背側部の広い範囲を占めるようになる。間脳は個体発生上、背側視床、腹側視床、視床下部および視床上部の四つの部位に分けられるが、その中で最も大きな部位を占めるのが背側視床である。単に視床といった場合は背側視床を指す。視床は第三脳室の両壁をなす卵円形の構造で、背側の遊離面は薄い髄質から成る帯層におおわれ、肺内側端に視床上部の構造である視床髄条が、前端より後方に走り手網核に付く。また背外側端は分界条によって終脳の尾状核と、外側方は外髄板によっておおわれ腹側視床の視床網様核と境されている。左右の視床は第三脳室内にまたがる視床間橋(中間質)によってつながり、視床下溝で視床下部と境される。視床の内部を構成している視床核は視床脚を介して大脳皮質と相互に結合する。内部には内髄板とよばれる線維板視床を内側部、外側部および前部に分けている。視床は感覚系と統合系との非常に重要な連絡部位である。嗅覚路以外のすべての感覚路がそれぞれ相当する視床の領域に投射する。「最近の研究によれば、嗅覚系も視床を投射する可能性がある」。視床で処理された感覚系情報の流れは視床大脳皮質線維を経て大脳皮質へと送られるが、大脳皮質の側からは多数の大脳皮質視床線維を介して視床における情報処理系に影響が及んでおり、したがって、視床と大脳皮質とは一つの機能単位としてはたらく。「運動」情報は小脳と大脳基底核を経て伝達され、統合系(大脳辺縁系や脳幹網様体など)からのさらに複雑な情報も視床に達する。したがって、視床は一方では大脳辺縁系と脳幹網様体との連結点として機能し、他方では大脳皮質も連絡しているわけである。
Zonal layer of thalamus(帯層;帯状層)Stratum zonale thalami たいそう;たいじょうそう
[A17_2_03_003]→
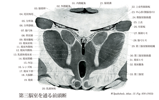
Thalamic nuclei(視床核;視床核群)Nuclei thalami
[A17_2_03_004]→視床は大きな灰白塊で、多くの細胞集団、すなわち視床核nuclei thalamiからなる。(解剖学講義)
Anterior nuclei of thalamus; Anterior thalamic nucleus(視床前核;前核)Nuclei anteriores thalami
[A17_2_03_005]→視床の前核群は視床の最も吻側で2分した内髄板によって囲まれた領域で背側表面の下にあり、ここに前結節を形成している。大きな主核である腹側前核(AV)と副核である背側前核(AD)および内側前核(AM)の3つの神経核が含まれる。ヒトでは腹側前核以外は退化的である。これらの諸核を構成している細胞は、中等大で、色素親和性の物質はほとんどなく、中等量の黄色色素が含まれ、有髄神経線維にとり囲まれている。乳頭体から線維を受け、帯状回と相互に線維連絡する。大脳辺縁系と関連しており、情動や新しい記憶と関連するという。
Anterodorsal nucleus of thalamus; Anterodorsal thalamic nucleus(前背側核)Nucleus anterodorsalis thalami
[A17_2_03_005_1]→前核群は乳頭視床束を受ける。乳頭体内側核からの線維は同側の前腹側核および前内側核に投射するが、他方、乳頭体外側核は、両側性に、前背側核に投射する。乳頭体外側核から出る神経線維は、背側前核と中脳被蓋の両者に両側性に投射する。
Ventral anterior nucleus of thalamus; VA(前腹側核)Nucleus ventralis anterior thalami
[A17_2_03_005_2]→VA核ともよばれる。前腹側核は、腹側核群の最吻側部に、そこでは、前方と腹外側方を網様核に境される。吻側方では、前腹側核が、視床のうち前核群の外側方の領域全体を占めるが、尾方においては、内側の領域の一部を占めるようになる。乳頭視床束は前腹側核を通過するが、尾方においては、内側の領域の一部を占めるようになる。乳頭体視床束は前腹束核を通過するが、その内側の境界をなしてはいない。この核は大形ないし中等大の多極細胞からなり、これらが集団をなして配列している。この核の中を太い有髄神経線維束が縦走する。この核のうち、乳頭視床束に隣接する部分およびこの核の腹方の境界に沿う部分は、大きな、暗調の細胞が密に配列し、大細胞部(VAmc)と呼ばれ、前腹側核の主部(VApc)よりも更に尾方に伸びる。このように、前腹側核には細胞学的に判然と区別される2つの部分があり、それぞれの部分は重複することなくいろいろ由来の異なる線維を受ける。
Anteromedial nucleus of thalamus; Anteromedial thalamic nucleus(内側前核;前内側核(視床の))Nucleus anteromedialis thalami
[A17_2_03_005_3]→前腹側核の内下方にある小さい核で大脳核、大脳皮質運動前野と連絡。錐体外路系と関連する。
Medial nuclei of thalamus(視床内側核;内側核)Nuclei mediales thalami
[A17_2_03_006]→視床内側核は視床内髄板と髄板内核に取り囲まれている。その内方および腹内方には視床正中線核群(midline thalamic nuclei)が分布する。正中線核群は菱形核、結合核、および紐傍核である。視床内側核は内側の巨大細胞部(magnocellular part)と外側の小細胞部(paraveocellular part)に区分される。小細胞部と内髄板の境界部に大型細胞から成る髄板傍部(pars paralemellaris)が記載されることがあるが、これはむしろ視床外側中心核(CL)に属するものと考えられる。視床内側核は腹側線条体からの投射もうける。この経路によって「側坐核(腹側線条体のもっとも主要な部分)-視床内側核-前頭前野皮質」の連絡系が成立する。視床から腹側線条体への投射線維は紐傍核(parataenial nucleus)から起こる。
Lateral thalamic nuclei; Lateral nuclei of thalamus(視床外側核;視床大核;視床外側核群)Nuclei laterales thalami; Nucleus magnus ししょうがいそくかく;ししょうたいかく;ししょうがいそくかくぐん
[A17_2_03_007]→視床前核群の尾方で視床の背外側部を占める。特定の上行性線維を受け連合核に属する。本核群は内髄板の背側縁に沿って広がる背側外側核(LD)とその尾方の後外側核(LP)とに分かれる。ヒトではLPの胞が大きく発達している。(医学書院医学大辞典:徳永叡)
Ventral anterolateral nucleus of thalamus(前外側腹側核)Nucleus ventralis anterolateralis [A17_2_03_007_1]→
Ventral intermediate nucleus of thalamus; VI(中間腹側核)Nucleus ventralis intermedius thalami
[A17_2_03_007_2]→視床の中間腹側核は腹側前核の後方にある核で、腹側核の中間1/3の複合体で、対側小脳(上小脳脚を通じて)、赤核と運動領皮質とのシナプスの中継核である。
Ventral posteromedial nucleus of thalamus; VPM(後内側腹側核)Nucleus ventralis posteromedialis thalami
[A17_2_03_007_3]→VPM核とも言われる。後内側腹側核は比較的淡く染まる神経網からなる三日月形の核で、後外側腹側核の内側で正中中心核の弯曲する境界の外側に位置する。後内側腹側核は次の明確な部分からなる。①小型、大型からなる主部で単に後内側腹側核とよばれる。②弓形の核の内側尖を占める小細胞性で淡染色の部で、小細胞部(VPMpc)とよばれる。後内側腹側核の主部は頭部、顔面、口腔内構造の受容期から体性求心性線維を受け、後外側腹側核の小細胞部は味覚に関係する。後内側腹側核の正確な境界はその線維結合に基づいて規定される。後内側腹側核は上行性二次賛辞神経線維を受けるがこれには、①内側毛帯に伴って上行する三叉神経脊髄路核と主感覚核からの交叉性線維、②三叉神経主感覚核の背側部からおこる背側三叉神経路の非交叉性線維が含まれる。ネコとサルの後内側腹側核からのHRP逆向性輸送により、対側の三叉神経核、特に中間部の尾側半、および三叉神経主核の腹側部の細胞が標識される。同側性に標識される細胞は大部分背側三叉神経路を生ずる主核の背内側部に限局する。顔面筋の伸展受容期からからのインパルスの一部も後内側腹側核の一部に両側性に伝達される。味覚を伝達する中枢経路はまだよくわかっていないが、味覚に関する感覚インパルスが後内側腹側核の内側小細胞部(VPMpc)に表現されることは確実である。鼓索、舌咽神経の電気刺激により、比較的短い潜時でVPMpcに誘発電位が示され体表面の触刺激にVPMpcは反応しないようである。ラットの味覚の表現は両側性といわれるが、ネコとサルではおもに同側性である。BenjaminとAlbert(Benjamin, R.M. 1959. Cortical and thalamic areas involved in taste discrimination in the albino rat. J. Comp. Neurol. 111: 231-260)は、ラットの皮質味覚野の切除によりVPMpcに限局した視床ニューロンの逆行性変性生じることを示した。動物実験の後内側腹側核の内側部の障害により味覚が失われる。ラットの脳神経の一次求心性線維により受容される味覚インパルスは、孤束核の吻側部に達し、そこで二次”橋味覚野”へ同側性に中継される。ラットの橋味覚野は、三叉神経中脳路と上小脳脚あいだの小細胞性の楔上部にある。この領域の細胞は上小脳脚の両側にあり、通常結合腕傍核とよばれる。これらの核の障害による変性は両側性に背内側被蓋を経てVPMpcへ追跡される。腹側前脳部へ達するといわれるこの橋味覚野からの側副路は、小窩講堂に影響する神経領域への直接の結合と考えられる。サルで迷走神経の入る部位より吻側の孤束核に注入されたアイソトープは、同側の中心被蓋路を経てVPMpcに直接達する上行線維束を標識する3H標識アミノ酸は、結合腕傍核とVPMpcに達する同側性線維束へ追跡された。VPMpcへのHRP注入により逆行性に、孤束核吻側部の外側亜分画のみが標識された。これらの所見は味覚が視床へ伝達される経路の理解を助ける。
Ventral posterolateral nucleus of thalamus; VPL(後外側腹側核)Nucleus ventralis posterolateralis thalami
[A17_2_03_007_4]→VPL核とも言われる。視床の後外側腹側核は腹側基底核群の外側部。内側毛帯と脊髄視床路の線維がシナプス結合する。内側毛帯線維の軸索終末の分枝は境界の明瞭な小円錐状を成して視床ニューロンとシナプス結合するが、脊髄視床路線維の軸索終末は散在性に分枝知る。入力情報はニューロンを乗り換えて大脳皮質の3、1、2野の下部へ投射する。
Lateral dorsal nucleus of thalamus; LD(背側外側核)Nucleus dorsalis lateralis
[A17_2_03_007_5]→LD核とも言われる。ラテン語をそのまま直訳すると外側背側核となるが、英語で用いられている背側外側核が一般的である。分類的には視床の外側核群-背側核-背外側核(LD)となる。中等大細胞からなり、特定の著明な上行性求心路をもたず、大脳半球の内側面の帯状回の後部および知覚中枢の後方における楔前部に投射する。このように一定の連合領域にひろく投射する核を連合核と呼び、これらはヒトでは特に発育がよく、外側核のほか後核や内側核がこれに属する。
Pulvinar of thalamus(後核;視床枕)Nucleus posterior; Pulvinar thalami
[A17_2_03_007_6]→視床枕は視床の後部と背外側を形成する大きな灰白質塊で、これの尾方は、内側膝状体、外側膝状体および中脳の背外側面の上に張り出している。視床枕は視床枕核群ともよばれており細胞学的にはかなり均一であるので、局所的な位置関係を基にして細分される。視床枕を形成するのは、明るく染まった、中等度の大きさの、多極性の細胞で、それらの細胞の分布密度と配列は、視床枕の部位によって異なっている。視床枕の前部の細胞は小さく、明るく染まり、散在性に配列している。下部は、視床枕の主部から、上丘腕の神経線維によって隔てられており、散在性の濃染する細胞で構成される。外側部を、外側髄板から広がる斜めの線維束が横切っている。視床枕の諸核は、長い、上行性の感覚神経路からは入力を受けてはいないが、その下部は、上丘の浅層のいくつかの層からの投射を受けている。局所的には、この投射は、反対側の視野の半分に相当する。視床枕の下部とそれい隣接する外側部とは、線条野を含む後頭葉の皮質と相互に結合する。視床枕の下部とそれに隣接する外側部はそれぞれ、反対側の視覚野の半分が復元され、網膜の部位局在的に、次の各部に投射している。すなわち、①皮質の18野と19野と、②有線野(17野)で、そこでは、線維が、顆粒層の上にある諸層に終止する。これらの結果によって、3つの視覚局在性をもった系統の入力が、視床(外側膝状体、視床枕の下部、およびそれに隣接する視床枕の外側部)から、一次視覚野に達し、しかも、そのいろいろな層に終止することが明らかになった。視床枕の下部から17野、18野および19野への投射は、膝状体外視覚神経路の中の最後の連絡を形成する。(視床枕の下核に隣接している部分以外の)視床枕の外側核は、側頭葉に投射し、同じ領域と相互に連絡している。視床枕の内側部は、上側頭回に投射しているらしい。
Reticular nucleus of thalamus(視床網様核;網様核(視床の))Nucleus reticularis thalami
[A17_2_03_008]→視床網様核は、外髄板と内包の間にある薄い神経核である。多くの神経線維の中に神経細胞が分布しているので網様核という名前が付けられているが、網様体とは関係がない。この核への求心性線維は、視床皮質線維と皮質視床線維の側枝である。網様核の神経細胞は、視床の神経核と相互連絡を持っており、この神経核の細胞の軸索は、自分に側枝をだしている視床の神経核に終止している。大脳皮質へは投射していない。
Medullary laminae of thalamus; Internal/external medullary lamina(視床髄板)Laminae medullares thalami; Lamina medullares [thalami] interna et externa
[A17_2_03_009]→視床髄板は視床の横断面に現れる有髄線維層。外側髄板は視床の腹側縁を形成し、視床下核や網様体核との境界をなす。内側髄板は視床の背内側核と腹側核との間を走り、髄板内核(中心内側核、中心傍核、外側中心核)を含む。
Intralaminar nuclei of thalamus(視床髄板内核;髄板内核)Nuclei intralaminares thalami
[A17_2_03_010]→視床髄板内核の特徴は、大脳皮質の広い範囲と線条体とに投射する点である。内髄板の中に分布している神経核を総称したもので、中心正中核(CM)、束傍核(Pf)、外側中心核(CL)、内側中心核(CeM)、中心傍核(Pc)が含まれる。中心正中核は大きな神経核で、背内側核と後内側腹側核の間にある。束傍核は、中心正中核の内側で、反屈束の近傍にある。髄板内核へは、網様体、青斑核、小脳、黒質、上丘、大脳基底核などの広い領域からの線維が入ってくる。脊髄視床路や三叉神経視床路も髄板内核に側枝をだしている。髄板内核からの線維は大脳皮質の広い範囲に投射しているほか、大脳基底核にも終止している。
Centromedian nucleus of thalamus (Centre médian de Luys); CM(中心内側核;中心正中核)Nucleus centromedianus thalami; Nucleus medialis centralis
[A17_2_03_011]→CM核とも言われる。視床の中心正中核は、髄板内核の中で最も大きくかつ、最も容易に認定される核であり、視床の中1/3部の高さにあり、背側内側核と後腹側核の間に位置する。その周囲は、内側の境界に沿って束傍核と融合する部分を除き、ほとんど完全に内側髄板の線維によって取り囲まれる。この核を構成する細胞は卵形ないし円形の細胞で、かなりの量の黄色色素(リポフスチン)を含む。この核の外側部にある細胞は小さいが、より内側方にあって背側内側核と境を接する領域にある細胞は、より大きく、かつ、いっそう密に配列している。網様体から線維を受け、大脳皮質に広く投射する。上行性網様体賦活系(大脳皮質を覚醒して意識を明晰に保つ)に関わる。
Medial geniculate nuclei(内側膝状体核)Nuclei corporis geniculati medialis
[A17_2_03_012]→内側膝状体核は聴覚情報の中継をなす細胞体群で、中脳の下丘からの線維も受け聴放線として側頭葉上部の聴覚皮質につながる。細胞群は腹側核、背側核、巨大細胞内側核に区分される。
Lateral geniculate nucleus(外側膝状体核)Nucleus cororis geniculati lateralis [A17_2_03_013]→
Habenular nuclei(手綱核)Nuclei habenulae
[A17_2_03_014]→手綱核は手綱の灰白質で、小型神経細胞からなる内側手綱核と大型神経細胞からなる外側手綱核からなる。両核は、脳基底部(中隔、基底核、外側視索前核)からの神経線維を受ける。さらに外側手綱核は淡蒼球の内節からの線維も受けている。両核は反屈束を通じて脚間核および中脳被蓋の内側域へ線維を出す。
Corpus striatum(線条体)Corpus striatum
[A17_2_03_015]→線条体は尾状核とレンズ核を意味する。レンズ核はさらに被殻と淡蒼球に区別される。このうち尾状核と被殻は終脳胞の腹外側に出現する神経節丘より同一の細胞群として発生し、その後、のちに発達してくる内包によって二つの部分に隔てられたものである。尾状核と被殻とは内包に横切って走る栓状の灰白質によって互いに連なり、特に前下方では両者は構造的にも同じ細胞構築をもっている。線条体という名称は内包を横切って尾状核と被殻を結んでいる灰白質によってできる縞目と、さらに、尾状核や被殻の中を走る有髄線維の小束によってできる縞目とに基づくものである。したがって、尾状核と被殻とをまとめて線条体Striatumとよび、淡蒼球をPallidumとよんで対比することが多い。解剖学用では、慣用されてきたStriatumという語とは異なる意味内容をもつ語として、Corpus striatumが採用されているが、日本名ではどちらも「線条体」である点は注意を要する。一方、淡蒼球の発生や細胞弧対句はStriatumとは異なる。淡蒼球は有髄神経線維に富むため黄灰白色を呈し、赤みを帯びた暗灰色のStriatumとは肉眼的にも明らかに識別できる。系統発生的視点に立って、尾状核と被殻を新線条体(Neostriatum)、淡蒼球を古線条体(Paleostriatum)、扁桃体を原線条体(Archistriatum)とよぶことがある。
Lentiform nucleus; Lenticular nucleus(レンズ核)Nucleus lentiformis
[A17_2_03_016]→レンズ核は小細胞性の被殻と大細胞性の淡蒼球を合わせたもので、その形が全体として両凸レンズに似ており、その全外面を神経線維群で包まれているため、一括してレンズ核とよばれる。尾状核頭および視床の腹外側にある大きい核で、前頭断でも水平断でも三角形で、頂点は内側に、底辺は外側にある。レンズ核は薄い外側髄板によって内側の淡蒼球と外側の被殻に分かたれる。レンズ核の内面は内包によって囲まれ、外面は外包によっておおわれる。しかし、両者の発生・細胞構築・線維連絡などは互いに異なる。被殻はレンズ核の外側面の近くにあって多少赤茶色を帯び、淡蒼球はレンズ核の内側面よりを占めて色も白っぽい。発生学的には、淡蒼球が最も古く、古線条体paleostriatumとも呼ばれる。これに対して尾状核と被殻は新しいので、この両者を合わせて新線条体neostriatum(または狭義の線条体striatum)という。また古線条体と新線条体をあわせたものすなわちレンズ核(淡蒼球+被殻)と尾状核を総称して、広義の線条体corpus striatumと呼ぶ(線状体ではない)。また広義の線条体にに扁桃体と前障を加えたものが大脳基底核basal gangliaである。大脳基底核の障害としては、パーキンソン病が有名である。
Putamen(被殻)Putamen
[A17_2_03_017]→被殻はレンズ核の外側部を形成し、外側髄板によって淡蒼球の外節とへだてられている。島皮質とは最外包、前障、外包によってわけられる。被殻の構造は尾状核とまったく同様で、太い有髄線維をほとんど含まず、主として小さい神経細胞からなるが、散在性の大細胞を含む。被殻と尾状核は発生学的にみると、同一の細胞群が内包の発達によって隔てられたもので、両者の間には互いに結合する灰白質の線条が多数見られる。そのため、両者をあわせて線条体または新線条体と呼ぶ。線維連絡も尾状核と原則的に等しい。霊長類において動物が高等になると、相対的な意味で尾状核の体積が減少し、被殻の体積が増大するといわれている。
Pallidum; Globus pallidus; Paleostriatum(淡蒼球;淡蒼部)Pallidum; Globus pallidus
[A17_2_03_018]→淡蒼球はレンズ核の最内側部を占め、被殻よりは小さい灰白質。垂直に走る板状の有髄神経線維(外側髄板)によって外側部の被殻と隔てられており、また、内側髄板によって内節と外節に分かれる。系統発生学的に線条体よりは古く、下等動物でよく発達している。発生に関しては、「間脳」性とするものののほか、その一部を「終脳」由来とするものがある。鉄反応が強陽性にでることが知られている。淡蒼球には大型の紡錘形ニューロンが多く、樹状突起は無棘で、長く髄板に平行して円板状に分枝する。淡蒼球の大型ニューロンの定量分析によれば、これらの細胞は単一のニューロン群に属する。淡蒼球の内節と外節の大型ニューロンには、なんら形態学的又は化学的な差異は見られない。ヒトでは外節は淡蒼球全体の約70%を占め、細胞密度が最も高い。淡蒼球の内節、外節の大型ニューロンはすべてGABA作働性である。淡蒼球ニューロンの軸索は少数の側枝を出している。淡蒼球の内側・外側髄板に存在する大型のコリン作働性ニューロンは、淡蒼球の腹側に存在する無名質のつづきである。多数の有髄線維束が淡蒼球を横走しているので、新鮮な脳では被殻や尾状核に比べ色が淡く見える。淡蒼球への求心性神経線維のもっとも主な起始は尾状核と被殻である。また、視床下核や黒質からの神経線維が内包を横切って主として内節に入る。淡蒼球が大脳皮質からの求心線維を受けるかどうかについては不確かである。視床からの求心線維については否定的な見解が多い。淡蒼球からおこる遠心性神経線維の主なものは視床と視床下核におわる。これらのうち、内節の腹側から出る神経線維は淡蒼球の腹側表面に集合してレンズ核ワナを形成し、内方後脚の腹内側部を背方にまわり、背尾側方へ走ってフォレル野に達する。一方、内節の背側からでる神経線維は淡蒼球の背内側部からおこる。これらは多数の小線維束として内包の腹側部を横切り、不確帯の腹側部に集合してレンズ核束(H2)を形成する。ついで、レンズ核束はフォレル野に入り、ここでレンズ核ワナの神経線維と一つになり、不確帯の内側端を取り囲むように腹側から背側にまわり、視床束(H1)を形成する。視床束は不確帯の背側を吻外方へ走り、視床に入って主として前腹側核(VA)、外腹側核(VL)、正中中心核(CM)などに分布する。なお、視床束には小脳視床線維も含まれている。淡蒼球から視床下核にいたる神経線維は主として外節からおこり、内包の尾側レベルにおいてその腹内側部を横切って視床下核に達する。淡蒼球からおこり中脳被蓋(脚橋被蓋核)や黒質へ向かう遠心性神経線維もある。
Caudate nucleus(尾状核)Nucleus caudatus
[A17_2_03_019]→尾状核は全体として弓状の大きな灰白質である。尾状核の吻側部は視床の吻側に位置し、側脳室前角のなかに隆起してその外側壁をなし、尾状核頭とよばれる。後方で細くなって尾状核尾とよばれ、視床の背外側縁に沿って側脳室の中心部の底面の外側縁を走り、やがて側脳室の弯曲に沿って下方にまがり、側脳室下核の上壁に達し、扁桃体との後端部のレベルでその外側部に接しておわる。尾状核頭と尾状核尾の中間部を尾状核体という。神経細胞には、大きく分けて、大小2種類(1:20)のものがみられる。求心性神経線維の起始部の主なものは、大脳皮質・視床髄板内核・視床正中中心核・黒質である。黒質(とくに緻密部)からはドパミン作働性の神経線維を受ける。尾状核からの遠心性神経線維の主な分布域は淡蒼球と黒質(とくに網様部)であり、これらの多くのものはGABA作働性である。
Amygdaloid nucleus(扁桃核)Nucleus amygdaloideus へんとうかく [A17_2_03_020]→
Claustrum(前障)Claustrum
[A17_2_03_021]→前障はレンズ核と島との間にある、内側が凹面をなす板状の核で、腹側方に厚くなる。この核とレンズ核との間には外包があり、また島の皮質との間には最外包がある。これらは狭い白質で、大部分は連合線維から、一部は交連および投射線維からなる。前障は種々の視床核、扁桃体などから線維を受け、大脳皮質に広く投射する。前障は以前は線条体とともにいわゆる基底核に数えられたり、あるいは皮質層の付け足しとして島皮質に属するものとされた。しかしながら、発生学的ならびに比較解剖学的研究によって、前障は発生の途中で位置がずれた古皮質の細胞群であることが証明されている。前障はその広い底の所で古皮質の領域へ移行する(すなわち梨状前野や扁桃体の外側核へ)。頭頂葉、側頭葉および後頭葉の皮質からの、無髄線維が局在的配列をなして前障に終わると言われている。前障の機能についてはわかっていない。
Hypothalamus(視床下部)Hypothalamus
[A17_2_03_022]→視床下部は間脳の中で、内臓機能、自律機能および内分泌機能と最も関係が深い部分である。これらの機能のすべてが、感情的および情動的行動と密接に関係している。第三脳室の側壁の下部および底にあたる。脳底面からみると、吻側から数えて、視交叉、漏斗、灰白隆起、乳頭体とつづき、漏斗の先端は下垂体に連なる。背側は視床下溝により視床と境されており、吻側は終脳の視索前野に、尾側は中脳被蓋と中脳中心灰白質に、尾外側は視床腹側部に移行する。通常、矢状面に平行な三つの帯状領域、すなわち、視床下部脳室周囲層、視床下部内側野、視床下部外側野に区分される。これらの間を多数の細かい神経線維が主として吻尾方向に走っている。視床下部は前後径が約10mmである。視床下部は内部環境を正常に維持する機序に関与し、また心悸亢進、瞳孔散大、“冷汗”の分泌などの情動反応の表出にも一役を演じている。大脳皮質を除去し、背側視床を除去した後でも、怒り反応はあらわれる(「みかけの怒りSham rage」)。さらに、視床下部は成長、性的成熟など身体の成熟過程にも関与している。したがって、視床下部を損傷すると、広汎で顕著な内分泌性、代謝性、行動性(情動性)の異常が一緒に起こってくることになる。視床下部は下垂体ホルモンによって内部環境に影響を及ぼすが、さらに脳幹網様体や自律神経系を介しても同様の働きを示す。視床下部への情報は通常の入力神経系によって伝達されるばかりでなく、視床下部のニューロンは内部環境からの物理的および化学的刺激(血液の温度など)にも反応する。植物性機能の中には、その統御中枢が視床下部自体に存在するものもあるが、呼吸や心臓の活動などの機能に関しては、視床下部はもっと下位の中枢に対する修飾器として働いている。
Supra-optic nucleus(視索上核)Nucleus supraopticus
[A17_2_03_023]→視索上核は正中線の両側にあり、その吻側部は視交叉の前上方に位置している。しかし、核の大部分は視索に沿って存在し、吻側野と尾側野に区分される。この核の構造は一様な中等大または大形(25~30μm)の神経細胞の集団で、視床下部内側野の神経核である。室傍核とともに代表的な神経分泌核あり、視索上核下垂体路の起始核である。
Paraventricular nucleus of hypothalamus(室傍核(視床下部の);視床下部室傍核)Nucleus paraventricularis hypothalami
[A17_2_03_024]→内側視床下野の吻側レベルでその背側部にある。視索上核とともに代表的な神経分泌核であり、室傍核下垂体路の起始核である。室傍核の細胞は第三脳室の上衣細胞層の直下にあり、細胞が密に集まって垂直な灰白質板を形成するが、その背側部は脳弓方向へと外側に伸びている。室傍核はいくつかの明瞭な細胞群で構成されていて、その細胞群の間には、内側部の主に小形の細胞群と顕著な外側の大型細胞群が存在する。
Ventromedial part of supra-optic nucleus(腹内側核;腹内側部(視索上核の))Pars ventromedialis nuclei supraoptici [A17_2_03_025]→
Ventromedial part of supra-optic nucleus(背内側核;腹内側部(視索上核の))Pars ventromedialis nuclei supraoptici [A17_2_03_026]→
Dorsal nucleus of hypothalamus(後核;背側核;視床下部背側核)Nucleus posterior hypothalami; Nucleus dorsalis hypothalami [A17_2_03_027]→
Lateral tuberal nuclei(隆起核;外側隆起核(視床下部の))Nuclei tuberales laterales りゅうきかく
[A17_2_03_028]→外側隆起核はヒトで最もよく発達しており、脳表面からもみえる外側隆起の直下において三つの長方形の水平な細胞柱を含む。中形の三角形のニューロンは容易には染色されない。この核の線維連絡はわかっていない。
Nuclei of mammillary body(乳頭体核)Nuclei corporis mamillaris にゅうとうたいかく
[A17_2_03_029]→後視床下部域にある細胞群で大型細胞からなる外側核さらに大きい内側核、背側核、腹側核、乳頭体上核で構成される。最初の2核は中脳の腹側に隆起して乳頭体を形成している。
Subthalamic nucleus(視床下核)Nucleus subthalamicus
[A17_2_03_030]→視床下核はルイ核ともよばれている。脳の断面の肉眼観察の際にも「目立つ」神経核であって、大型ニューロンから成り、間脳の最尾部において内包後脚の背内方に位置している。この神経核の内側部は黒質吻側部の背方に位置する。背側の不確帯とはレンズ核束(H2)によりへだてられている。核の境界は明瞭で、前頭断面では両凸レンズ形を呈し、矢状断面ではほぼ円形を呈する。核の尾側端のレベルでは、核の内側部が黒質の最吻側端の背縁に接している。主な求心出力神経線維を淡蒼球や脚橋被蓋核受け、また、遠心性神経線維を主として淡蒼球内節に送る。大脳皮質とくに前頭葉からの求心性線維や、黒質や淡蒼球への遠心性線維の存在が報告されているが、その他の線維連絡関係については不確実な点が多い。ヒトでこの核が損傷されると、反対側の半身に激しい不随意運動、すなわちヘミバリスムがおこる。視床下核の細胞はグルタミン酸塩を含有しており、淡蒼球と黒質のニューロンに興奮性に作用すると言われている。グルタミン酸塩は細胞の基礎代謝にも存在するもので、視床下核の細胞がグルタミン酸塩免疫反応陽性であっても、かならずしもグルタミン酸塩がこの核の細胞により使用される神経伝達物質とはいえない。視床下核は視床下部外側核の最後の細胞集団から発生してくる。吻側の細胞集団は淡蒼球の内節、外節の原基となる。Luys, Jules Bernard (1828-1898)フランスの神経学者。
Interpeduncular nucleus(脚間核)Nucleus interpeduncularis
[A17_2_03_031]→脚間核は脚間窩の背側にある。この核は手綱核からの投射を反屈束を経由し受けている。多くのコリン作働性線維が脚間核に終止する。反屈束の線維のうちで脚間核を通過するものは上中心核、背側被蓋核および中脳中心灰白質に達している。背側被蓋核はまた乳頭体被蓋路を通して乳頭体からも線維を受け、中脳中心灰白質の腹内側にある小さな線維束である背側縦束と密接に関係している。これらの神経路は辺縁系に関連した信号を中脳の諸核に伝えている複雑な神経路の一部を構成しており、内臓や行動機能に関係する。
Mesencephalon; Midbrain(中脳)Mesencephalon
[A17_2_04_001]→中脳は「中央」を意味するギリシャ語の接頭詞mesoと、「脳」を意味するencephalonを結合したもの。中脳、橋、延髄を合わせて脳幹と呼ぶが、これは頭蓋底の大後頭孔の所から上方に向かい、大脳半球の基底面にまで伸びる楔形の構造である。中脳は狭義の脳幹の最上方部で、上方に間脳、下方に橋との間の中脳水道を囲む比較的上下に短い構造を指す。間脳との境は厳密には不明確であるが、背側に後交連の後部、腹側に乳頭体の後方を通る面で境される。下方は背側に下丘の後方と腹側の橋の前方を通る面で比較的明確に境される。外形を見ると背側に蓋板によって形成された4個の隆起があり上方の一対を上丘、下方の一対を下丘という。上丘および下丘からは上外側に線維束を出し、それぞれ上丘腕および下丘腕として間脳につづく。腹側には大脳脚がみられ、その間に多数の小血管が通る後有孔質の間の細い溝を大脳脚内側溝とよび、ここから動眼神経の根がでる。断面では背側部は蓋板で包まれ、(視蓋とも呼ばれる)上丘および下丘を形成し、その腹側端はほぼ中脳水道の中央部を通る面で区切られる。これより腹側を広義の大脳脚というが、これはさらに中脳被蓋と狭義の大脳脚にわけられる。中脳では固有の細胞集団と線維束があり、細胞群としては中脳水道を取り囲む中心灰白質が三叉神経中脳路および核によって外側を包まれ、腹側正中部には上方に動眼神経核、下方に滑車神経核が存在する。また上方の動眼神経核の腹外側に赤核があり、さらに腹側に大脳脚の背側面を覆って黒質が存在する。正中腹側端部の大脳脚にはさまれた部位には脚間核がある。中脳に出入りする線維束で著明なものは中心灰白質内には腹外側部に背側縦束があり、赤核の背側および背外側方に中心被蓋路がある。さらに上丘中央から下丘の高さで正中部に強い線維の交叉がみられる。これらの交叉は被蓋交叉および上小脳脚交叉で、被蓋交叉の背側部は多くは上丘および上小脳脚交叉で、被蓋交叉の背側部は多くは上丘および被蓋からの下行線維から成り、腹側部は赤核からの下行線維から成る。また上小脳脚交叉は小脳核から赤核および視床へ投射する線維の交叉部である。
Red nucleus(赤核)Nucleus ruber
[A17_2_04_002]→赤核は中脳被蓋では最も明瞭な部分で、網様体のほぼ中央にあり、桃黄色を呈し、周囲を上小脳脚線維で囲まれている。赤核全体として卵円形で、横断面は円形を呈し、上丘の下端から間脳の下部まで広がる。細胞学的には後方の大細胞部と前方の小細胞部によりなる。赤核の細胞の間には主として上小脳脚からの有髄線維の細い束が存在する。動眼神経根の一部が脚間窩に至る途中で赤核を貫く。赤核の求心性線維は主として小脳核および大脳皮質からくる。これらの線維は体部的局在性をもって赤核に終止する。上小脳脚の線維は中脳下部で完全に交叉し、対側の赤核およびその周囲に至る。小脳歯状核からの側枝は対側赤核の上1/3部に終止し、一方、栓状核(前中位核)および球状核(後中位核)からの線維は赤核の下2/3部に体部位的局在性をもって終わる。この連絡は小脳の傍虫部皮質と赤核大細胞部をつなぎ、さらに赤核から脊髄へ体部位的局在性に投射する。これには上小脳脚交叉および赤核脊髄路の交叉の2つの交叉が含まれる。赤核の大細胞部からの体部位的局在投射は主に頚髄と腰髄に終わる。皮質赤核線維は中心前回や前運動野から起こり、ともに両側の赤核小細胞部に体部位的局在性をもって終止する。大脳皮質6野の内側部(補足運動野)からの投射線維は対側の赤核大細胞部に終わる。中心前回の運動野から赤核大細胞部に終止する線維は同側性で、赤核運動路の体部位的局在性をもつ起始に対応する。これらの皮質赤核線維は体部位的局在性をもち、赤核脊髄路(交叉性)と共に運動野皮質から脊髄へインパルスを伝える経路の一部をなす。赤核からの下行性遠心路は腹側被蓋交叉で交叉し、①小脳の中位核、②三叉神経主知覚核および脊髄路核、③顔面神経核の一部、④いくつかの延髄の中継核および⑤脊髄に投射する。また赤核の小細胞部からの非交叉性の線維束が中心被蓋路に入り、オリーブ核主核の背側板に終わる。これを赤核オリーブ路といい、小脳へのフィードバック系の一部をなす。赤核からの視床への投射はない。
Substantia nigra(黒質;黒核)Substantia nigra; Nucleus niger
[A17_2_04_003]→黒質は中脳被蓋腹側部の核で大脳脚の背側に接して存在する。ヒトの黒質の神経細胞は顆粒状のメラニン色素を豊富に含有するため、黒質は全体として肉眼的に黒くみえる。黒質には背側の緻密部と腹側の網様部が区分される。緻密部が神経細胞に富むのに対し、網様部では神経細胞の密度は粗で、細い神経線維に富む。したがて、前者は黒色部、後者は赤色部とよばれることがある。黒質からおこる遠心性神経線維としては、緻密部からおこり線条体に分布する黒質線条体線維、網様部から起こり視床のとくに内側腹側核(VM)に分布する黒質視床線維、および網様部からおこり上丘の中間灰白質に分布する黒質上丘線維などが主なものである。また、黒質に分布する求心性神経線維の起始としては、線条体・淡蒼球・視床下核(Luys体)が主なものである。これらのほか、前頭葉皮質・背側縫線核・扁桃体中心核・外側手綱核なども報告されているが不確実である。黒質は中枢神経系のうちでドーパミンとGABAの含有量が高い部位として知られる。ドーパミンは線条体に神経線維を送る黒質緻密部の神経細胞に主として含まれ、またGABAは線条体よりおこり黒質網様体に至る神経線維の軸索終末に主として含まれる。黒質に見られる線維はまたは11個のアミノ酸が連絡したペプチドとしてのP物質(SP)も含む。黒質は脳において最も高濃度にP物質を有する部位で、この物質は黒質の緻密部および緻密部内の神経終末に凝集している。網様部はまたエンケファリン作働性線維および終末も有する。尾状核および被殻の樹状突起の棘突起に含むニューロンから起こる線条体黒質線維はGABA、P物質、エンケファリンを含む。これらの線維は同様の伝達物質を有する線条体淡蒼球線維を出すニューロンとは異なる細胞集団から起こる。黒質はパーキンソン病(振戦麻痺)の原因となっている代謝障害に緻密に関係しており、Huntington舞踏病および異常な不随意運動や筋緊張の変化を特徴とする他のタイプの運動障害にも関与しているようである。パーキンソン病では黒質から線条体へのドーパミンの輸送および合成が極度に傷害される。Huntington舞踏病では線条体のドーパミンは正常であるがGABAは著明に減少している。
Central gray substance(中心灰白質;中心灰白層)Substantia grisea centralis; Stratum griseum centrale hypothalami ちゅうしんかいはくしつ
[A17_2_04_004]→中心灰白質は一般用語としては、脊髄の中心管と脳幹の第3脳室、第4脳室に隣接する、またはそれらを取り巻く、主に小形の細胞からなる灰白質をいう。特殊用語としては中脳水道を取り巻く灰白質の厚い袖で、吻方で視床下部の後核に連続しているもをいう。
Reticular formation(網様体)Formatio reticularis
[A17_2_04_005]→網様体は延髄、橋、中脳においてその中心をなす構造で、系統発生的に古く、あまり境界の明瞭でない細胞群(核)からなっている。核を構成する細胞は多極性で、その形、大きさも変化に富んでいる。樹状突起は長く、広汎にに分岐し、そこに多種類の入力が収束する。軸索は上行枝と下行しに二分し、豊富な側枝により複雑な結合を行う。細胞構築学的には次の三つのあての細胞柱が区別される。①縫線核と正中傍網様体核群、②内側核群(腹側網様体核、正中網様体核、巨大細胞網様体核、橋被蓋網様体核)、③外側群(外側網様体核、小細胞網様体核、楔状核、脚橋被蓋核)。これらの核は種々の経路を介して非常に広汎な領域と関係している。入力は脊髄(脊髄網様体路、脊髄視床路)、脳神経(Ⅴ、Ⅶ、Ⅷ、Ⅸ、Ⅹ脳神経の中枢枝)、大脳皮質(皮質網様体路)、小脳(室頂核網様体路、歯状核網様体路)、辺縁系(内側前脳束、背側縦束)、中脳(視蓋網様体路)に由来する。出力は視床(網様体視床路[中心被蓋路])、小脳(外側、正中傍、橋被蓋網様体核からの網様体小脳路)、脊髄(網様体脊髄路)などにいたる。また網様体にはモノアミン含有細胞が多数存在し、中枢神経系内で広汎な結合を行っている。縫線核群は世路と飲を含有する。核により結合は異なるが、脊髄脳神経核(Ⅴ-Ⅷ脳神経核、迷走神経背側核、弧束核)、オリーブ核、中心灰白質、小脳、四丘体、黒質、間質核、Darkschewitsch核、視床、視床下部、前頭葉、扁桃核、梨状葉、嗅球、嗅結節、大脳基底核などに線維を送る。アドレナリン含有細胞は青斑核のほかに弧延髄被蓋の腹側部や背内側部(孤束核の腹内側)に存在する。これらの細胞も広く、脊髄、脳神経核(ⅩⅡ、Ⅶ、Ⅴ脳神経核、迷走神経背側核、孤束核、蝸牛神経核)、オリーブ核、中心灰白質、四丘体、視床、視床下部、海馬、レンズ核、大脳皮質などに線維を送っている。
Tegmental nuclei; Nucleus of Gudden(被蓋核)Nuclei tegmenti (tegmentales); Nucleus tegmenti dorsalis et ventralis ひがいかく [A17_2_04_006]→
Interstitial nucleus(間質核)Nucleus interstitialis
[A17_2_04_007]→間質核は内側縦束の傍らにあり、比較的大きい多極性の細胞からなる。この核から出る線維は後交連を通って正中線で交叉し、動眼神経核、滑車神経核、前庭神経内側核などとシナプス結合する。これらの線維が障害されると垂直方向の眼球麻痺が起こる。
Gray layer of superior colliculus(上丘灰白層)Stratum griseum colliculi superioris じょうきゅうかいはくそう [A17_2_04_008]→
Pretectal nucleus(視蓋前核)Nucleus pretectais しがいぜんかく
[A17_2_04_009]→
Nucleus of trochlear nerve; Trochlear nucleus(下丘核)Nucleus nervi trochlearis
[A17_2_04_010]→滑車神経核は下丘の高さで、中心灰白質の腹内側にあり、動眼神経核の直接尾方延長部にあたる。これは動眼神経主核と同様に多極性の大細胞からなり、これから出る根線維は中心灰白質の外側部を背側方でやや下方に走り、下丘の下で完全交叉をおこない(滑車神経交叉)、脳を出る。なお滑車神経核の背側で中心灰白質中には背側被蓋核が認められ、また内側縦束の腹側には腹側被蓋核が区別される。
Nucleus of oculomotor nerve(動眼神経核)Nucleus nervi oculomotorii
[A17_2_04_011]→動眼神経核は第三脳神経、すなわち、動眼神経の起始核であり、動眼神経運動神経細胞の集合である。中脳被蓋の正中線背側部の両側に存在し、中脳中心灰白質の腹方に位置する。動眼神経核の尾側端は滑車神経核の吻側端とほとんど連続しており、動眼神経核の吻側端は中脳の最吻側レベルに達する。動眼神経運動神経細胞はそれぞれの筋支配に対応して局在配列する。すなわち、下直筋支配細胞は核の背側部、内直筋支配細胞は核の腹側部、下斜筋支配細胞は前2者の中間部、上直筋支配細胞は内側部に位置し、それぞれ脳幹の長軸に平行な左右一対の細胞柱を形成する。これらのうち、上直筋支配細胞柱だけは反対側の筋を支配するが、その他は同側の筋を支配する。また、上眼瞼挙筋を支配する細胞柱は正中部に位置し、不対であって、動眼神経核の尾側1/3のレベルにのみ存在する(尾側正中核)。以上の他、副核(Edinger-Westphal核)および正中核(Perlia核)を含めて動眼神経核群とされることがおおい。
Nucleus of trochlear nerve; Trochlear nucleus(滑車神経核)Nucleus nervi trochlearis
[A17_2_04_012]→滑車神経核は下丘の高さで、中心灰白質の腹内側にあり、動眼神経核の直接尾方延長部にあたる。これは動眼神経主核と同様に多極性の大細胞からなり、これから出る根線維は中心灰白質の外側部を背側方でやや下方に走り、下丘の下で完全交叉をおこない(滑車神経交叉)、脳を出る。なお滑車神経核の背側で中心灰白質中には背側被蓋核が認められ、また内側縦束の腹側には腹側被蓋核が区別される。
Mesencephalic nucleus of trigeminal nerve(三叉神経中脳路核;三叉神経中脳核)Nucleus mesencephalicus nervi trigeminalis
[A17_2_04_013]→三叉神経中脳路核は三叉神経運動核の下端より少し下方の高さから中脳上端部に至る。非常に細長く延びた核で、橋の高さでは第四脳室の腹外側核の近くにあり、中脳では中心灰白質の外側縁にある。これは脊髄神経節の細胞に似た、少数の大きい偽単極細胞からなる。その突起は三叉神経中脳路を作りつつ下行し、三叉神経運動核に突起を出したのち運動根に加わり、主として咀嚼筋に分布し、その固有知覚を伝え、咬む力の調節に関与している。また中脳路核上部には外眼筋などからの固有知覚が伝えられるという。中脳路核から高次の中枢への連絡については不明である。
Lateral mesencephalic ganglion; Lateral nucleus of mesencephalon(外側中脳神経節;中脳外側核;中脳背側核)Ganglion mesencephali laterale; Nucleus laterale mesencephali; Nucleus dorsalis mesencephali がいそくちゅうのうしんけいせつ;ちゅうのうがいそくかく;ちゅうのうはいそくかく
[A17_2_04_013_1]→
Pons(橋)Pons
[A17_2_05_001]→Ponsとは、橋(ハシ)という意味である。腹側から見ると左右の小脳半球の間に架かった太鼓橋の様に見えるところから橋という名前が付けられた。比較解剖学的には、橋が延髄から区別されるのは哺乳類に限られ、橋は人類で最もよく発達している。後脳の腹側部にあたる。すなわち、小脳の腹側に位置しており、延髄と中脳の間に介在する。橋の腹側面は横走する幅広い神経線維束(横橋線維)によっておおわれる。この神経線維束はさらに橋の外側面において、橋と小脳を連結する中小脳脚を形成しており、左右の小脳半球の間にかかる「橋」のようにみえる。橋は既にユースタキウスEustachius (1524-1574)の図に載っているというが、この図は1714年まで出版されなかったので、Ponsという名称は、このような外見に基づいて、イタリアの解剖学者であり外科医でもあったC.Varolio (1543-1573)が用いたものである(ヴォロイオ橋)。橋は横断面では橋腹側部または橋底部と橋背部または橋被蓋とに区分される。両者の境界は橋被蓋の腹側部を上行する内側毛帯の腹側縁にあたる。橋底部の神経線維群には、上記の横橋線維のほかに、橋底部の中心部を縦走する橋縦束があり、神経細胞としては橋縦束を取り囲んで橋核が存在する。橋縦束の線維はその大部分が大脳皮質からの下行神経線維であり、橋核に連絡する皮質橋核路を含む。橋核は大脳皮質からおこる求心性神経線維のほか、小脳核や上丘からおこる求心性神経線維を受けることが知られている。橋核からおこる遠心性神経線維は横橋線維、ついで中小脳脚を形成して、主として反対側の小脳半球の皮質に連絡する。また、その際、小脳核、とくに歯状核に側枝を送る可能性が大きい。このように、橋縦束・橋核・橋横線維は大脳皮質や小脳半球など、系統発生的に新しい部位との関係が深く、哺乳動物ではじめて出現する構造であって、高等な哺乳類において良好な発育を示す。 一方、橋被蓋は系統発生的に古い構造であり、脳幹網様体の基本構造を示す部位がもっとも広い領域を占める。脳神経核としては、三叉神経核(主感覚核・脊髄路核・中脳路核・運動核)・顔面神経核・内耳神経核(蝸牛神経核と前庭神経核)が存在する。また、橋被蓋の外側部を上行する外側毛帯、および橋被蓋の腹側部を横走する台形体の線維は聴覚路を形成する神経線維群であり、聴覚神経路の中継核として、外側毛帯核および台形体核が存在する。その他の線維群としては、第四脳室底の腹側において正中線背側部の両側を内側縦束が縦走し、上小脳脚が第四脳室蓋の外側部を形成している。また、神経細胞群としては、橋被蓋の背外側部に青斑核が、上小脳脚の周辺部には結合腕傍核が存在する。
Motor nucleus of facial nerve(三叉神経核)Nucleus nervi facialis
[A17_2_05_002]→三叉神経核は咀嚼筋を支配している運動核と主にその固有感覚に関与する中脳路核、および顔面、口腔内領域と脳膜の一部の一般体性感覚に関与する感覚核群からなる。感覚核群はさらに主感覚核および脊髄路核からなる。運動核は橋の腹外側から脳幹を出て感覚根の内側部に沿って走り、三叉神経節の下部を通って咀嚼筋にいたる。感覚核群のうち主感覚核は橋の高さで運動核の背外側方に位置して存在する。主感覚核より下方につづいて脊髄路核が存在するが、これは延髄の全長にわたり背外側に位置し、下方に頚髄第2節の高さにまで伸び、次第に頚髄後核に移行する。この核は細胞構築的に上方より上位核、中位核および下核に細分される。感覚核群は主感覚核および脊髄路核ともに三叉神経節(半月神経節)の中枢枝の線維を受ける。脊髄路核にいたるこれらの線維は、核の外側方をおおって三叉神経脊髄路を形成している。三叉神経脊髄路内では末梢の三枝からの局在が明確で、腹側より眼枝、上顎枝および下顎枝からの線維がその順に背側方に向けて並ぶ。さらに脊髄路の背内側端には舌咽および迷走神経の中枢枝が加わる。感覚群からは第2次経路としてそれぞれの高さから視床に向かう線維がでる。主感覚核の背側部から出た線維は中心被蓋路に沿って上行枝の視床後内腹側核におわる。主感覚核の腹側部および脊髄路核から視床へ向かう線維が出る。主感覚核の背側部からでた線維は中心被蓋路に沿って上行し同側の視床後内腹側核におわる。主感覚核の腹側部および脊髄路核から視床へ向かう線維はそれぞれの高さで交叉し、対側の内側毛帯と併走して三叉神経毛帯をなして対側の視床後内側腹側核にいたる。上記の視床にいたる線維のほかに、感覚核群からは両側の網様体、対側の上丘および小脳に線維を送る。機能的には主感覚核は主に触圧覚に関与し、脊髄路核ことに下核は温痛核に関与するといわれる。 第7脳神経、すなわち顔面[中間]神経を形成する神経線維のうち、表情筋・広頚筋・アブミ骨筋・茎突舌骨筋などの横紋筋を支配する運動神経線維の起始核であり、橋の最尾側レベルにおいて橋被蓋の腹外側部に位置する。顔面神経核からおこる神経線維は核の背側から出て背内頭側に走り(顔面神経上行根)、第四脳室底の直下で外転神経核の内側部に達してはじめて密な神経束を形成する。ついで、この線維束は外転神経核の頭側レベルで核の背側を外側に向かい顔面神経膝を形成する。ついで、線維束は三叉神経脊髄路核の内側縁に沿うように腹外側に走り(顔面神経下行根)、橋の尾側レベルで脳幹を出る。顔面神経の支配を受ける横紋筋のうち顎二腹筋後腹は副顔面神経核に支配される。副顔面神経核の神経細胞は顔面神経核と三叉神経運動核を結ぶ線上に散在性に存在する。顔面神経に含まれる副交感神経線維の起始核として、上唾液核が記載されている。この核の神経細胞は、橋被蓋網様体の尾側レベルでその背外側部において、三叉神経脊髄路核の内側縁付近に比較的散在性に存在するようである。
Nucleus of abducens nerve; Abducens nucleus (VI)(外転神経核)Nucleus nervi abducentis
[A17_2_05_003]→第6脳神経、すなわち、外転神経の起始核で、眼筋のうち外側直筋を支配する運動神経細胞群である。第四脳室底において橋の正中線背側核の両側に位置しており、顔面神経膝とともに、菱形窩に低い隆まりを形成する。
Nuclei of solitary tract; Solitary nuclei(孤束核)Nuclei tractus solitarii
[A17_2_05_004]→孤束核は延髄背側を矢状方向に走る細い細胞柱で、菱形窩底の下で境界溝のすぐ外側にある。小さい独立核の集まりで一括して言えば脳幹の内臓求心性の核であって迷走神経、舌咽神経、顔面神経からの線維が孤束を経てはいってくる。孤束核は次の数個の部分に分けられる。すなわち、①内側部、迷走神経背側運動核の背外側にある、②背内側、背外側および腹外側亜核、孤束を取り巻く、および③小細胞性亜核、最後野の腹側にある。内側部の細胞は迷走神経背側運動核を吻側にわずかに越え、また、第四脳室より下方へ広がり、反対側の同じ細胞性と合一して迷走神経交連核をつくる。外側の諸核は大細胞性の細胞柱をなし、孤束を一部あるいは完全に取り巻いている。この外側部は孤束のほぼ全長にわたって平行に存在する。吻側では橋下部まで広がり、一方尾側ではその細胞数が減少し、網様体との区別が難しくなる。孤束核の膨大した吻側部(すなわち外側部)は主として特殊内臓求心性(味覚)線維を顔面神経(中間神経)と一部の顔面神経と舌咽神経からくる一般内臓求心性線維を受ける。孤束核の内側部の細胞は多数のニューロペプチドを含有している。すなわち、エンケファリン、ソマトスタチン、サブスタンスP、およびコレシストキニンなどである。同部位にはサブスタンスP含有線維も豊富に存在する。孤束核から起こる二次路系は次の部位に同側性に投射する。すなわち、①疑核とその周辺の網様体、②橋上部の傍腕核、および③味覚に関与する視床の核、すなわち後内側腹側核(小細胞部)である。孤束核から起こるその他の二次線には舌下神経核と唾液核に投射し、舌の運動と唾液の分泌反射を仲介する。孤束核からの迷走神経背側運動核、横隔神経核(第三(C3)・第四(C4)・第五(C5)頚髄の高さ)、および胸髄の前角細胞への投射はせきと嘔吐反射に関与する。孤束核は生理学的に同定された延髄の呼吸“中枢”と同一の広がりをもち、この中枢は疑核とその周辺の網様体を包含する。延髄“呼吸中枢”の細胞は迷走神経のインパルス、および直接の化学的環境変化(CO2蓄積)によって賦活される。別個の昇圧帯と減圧帯からなる延髄の血管運動“中枢”は十分明らかにされていない。最近、心臓血管系の調節に関する神経回路網が重視されてきた。ノルアドレナリン作働性ニューロンの1集団(グループA5と呼ばれる)が橋の下部で上オリーブ核と顔面神経の根線維の間にあり、軸索を孤束核、疑核、迷走神経背側運動核および胸髄の交感神経節前線維のニューロンに投射しており、上記神経回路網の一部と思われる。
Nucleus of vestibulocochlear nerve(内耳神経核)Nuclei nervi vestibulocochlearis
[A17_2_05_005]→内耳神経核は菱形窩の中央部の外側部から下方にわたって存在し、前庭神経核と蝸牛神経核とに分けられる。(解剖学講義)
Nuclei of trapezoid body(台形体核)Nuclei corporis trapezoidei
[A17_2_05_006]→台形体核は台形体の線維の間に散在する細胞よりなり、台形体の線維や聴条の線維がこれとシナプス結合する。台形体外側核、内側核、前(腹側)核がある。聴覚の中継核とみられる。ここからの線維の大部分は外側方に走り、外側毛帯に入るが、その近くに上オリーブ核群と呼ぶ大きな神経細胞集団がある。
Anterior nucleus of trapezoid body; Ventral nucleus of trapezoid body(台形体腹側核)Nucleus anterior corporis trapezoidei
[A17_2_05_007]→台形体の外側部で、その線維のあいだに散在する。
Posterior nucleus of trapezoid body(台形体後核;台形体背側核;後脳オリーブ核)Nucleus corporis trapezoidei posterior; Nucleus olivaris metencephali だいけいたいこうかく;だいけいたいはいそくかく;こうのうおりーぶかく
[A17_2_05_008]→
Nuclei of lateral lemniscus(外側毛帯核;外側絨帯核)Nuclei lemnisci lateralis
[A17_2_05_009]→外側毛帯核は背腹方向に長い帯状の神経核で、橋被蓋を中脳に向かって上行する外側毛帯中にある核で、外側毛帯の線維のあいだにはニューロンが散在している。腹側核、中間核、背側核に区別される。外側毛帯の線維の中にはこれらのニューロンとシナプス結合するものがある。左右の外側毛帯背側核と腹側核の間には、プローブスト交連Probst's commissureとよばれる交連線維がある。外側毛帯を構成する線維は上行し、下丘に終わっている。外側毛帯は脳幹の聴覚路の中でもっとも重要な物である。
Pontine nuclei(橋核)Nuclei pontis
[A17_2_05_010]→橋核は橋腹側部中にある核で小脳前核(脳幹から小脳皮質への求心線維は、大部分が下オリーブ核や延髄にある多くの小さな核で中継されるが、このようにほとんどの線維を小脳に送るような脳幹の核を小脳前核と総称する)としては最大のもので、かつ大脳皮質からの興奮を小脳に伝えるもっとも重要な中継核である。皮質橋路線維は大脳の各葉から起こり、同側の橋核に終止する。一次運動野、感覚野および視覚野の一部からの線維が大部分をしめる。運動野と体性感覚野からの投射はそれぞれ体部位的局在をもって配列するが、その終枝部位は別々である。すべての橋核からの線維は中小脳脚を通るが、小脳半球部皮質至る物が対側性であるのに反し、中部皮質へ至るものは両側性である。小脳小節だけは橋核からの投射がない。橋小脳線維は帯状線維として終止し、小脳の各小葉は大部分橋核内の2カ所以上の異なった部位からの投射をうける。
Reticular formation(網様体;橋網様体)Formatio reticularis
[A17_2_05_011]→網様体は延髄、橋、中脳においてその中心をなす構造で、系統発生的に古く、あまり境界の明瞭でない細胞群(核)からなっている。核を構成する細胞は多極性で、その形、大きさも変化に富んでいる。樹状突起は長く、広汎にに分岐し、そこに多種類の入力が収束する。軸索は上行枝と下行しに二分し、豊富な側枝により複雑な結合を行う。細胞構築学的には次の三つのあての細胞柱が区別される。①縫線核と正中傍網様体核群、②内側核群(腹側網様体核、正中網様体核、巨大細胞網様体核、橋被蓋網様体核)、③外側群(外側網様体核、小細胞網様体核、楔状核、脚橋被蓋核)。これらの核は種々の経路を介して非常に広汎な領域と関係している。入力は脊髄(脊髄網様体路、脊髄視床路)、脳神経(Ⅴ、Ⅶ、Ⅷ、Ⅸ、Ⅹ脳神経の中枢枝)、大脳皮質(皮質網様体路)、小脳(室頂核網様体路、歯状核網様体路)、辺縁系(内側前脳束、背側縦束)、中脳(視蓋網様体路)に由来する。出力は視床(網様体視床路[中心被蓋路])、小脳(外側、正中傍、橋被蓋網様体核からの網様体小脳路)、脊髄(網様体脊髄路)などにいたる。また網様体にはモノアミン含有細胞が多数存在し、中枢神経系内で広汎な結合を行っている。縫線核群は世路と飲を含有する。核により結合は異なるが、脊髄脳神経核(Ⅴ-Ⅷ脳神経核、迷走神経背側核、弧束核)、オリーブ核、中心灰白質、小脳、四丘体、黒質、間質核、Darkschewitsch核、視床、視床下部、前頭葉、扁桃核、梨状葉、嗅球、嗅結節、大脳基底核などに線維を送る。アドレナリン含有細胞は青斑核のほかに弧延髄被蓋の腹側部や背内側部(孤束核の腹内側)に存在する。これらの細胞も広く、脊髄、脳神経核(ⅩⅡ、Ⅶ、Ⅴ脳神経核、迷走神経背側核、孤束核、蝸牛神経核)、オリーブ核、中心灰白質、四丘体、視床、視床下部、海馬、レンズ核、大脳皮質などに線維を送っている。
Reticular formation of pons(網様体(橋被蓋の);橋網様体)Formatio reticularis pontis きょうひがいのもうようたい;きょうもうようたい
[A17_2_05_011]→橋網様体は、主として下(橋)網様核と上(核)網様核の2つの大きな細胞集団よりなる。下網様核は延髄の巨大細胞網様核の上方部に相当し、上方は三叉神経運動核の高さにまで及ぶ。上網様核は、上方が中脳下部にまで伸びるが、正確な境界は明らかではない。橋網様体の細胞からは非交叉性の網様体脊髄路が起こり、脳幹では内側縦束の一部として下行する。その他の細胞からの線維には中心被蓋路の一部として上行するものもあり、また多くの細胞の線維は二分して上方および下方に分枝を送る。このうち上行枝は中心被蓋路を経て視床の髄板内核に投射する。これらの視床核に至るインパルスは大脳皮質の広い部位の電気活動に強い影響を及ぼす。橋にあるその他の網様核として被蓋網様核と上中心核がある。前者は縫線の近くで内側毛帯の背側にあり、胸郭が被蓋の中にあり、菱脳峡の高さで大きくなり、縫線正中核となる。
Substantia ferruginea(鉄色質)Substantia ferruginea
[A17_2_05_012]→
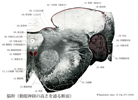
Myelencephalon; Medulla oblongata; Bulb(延髄)Myelencephalon; Medulla oblongata; Bulbus
[A17_2_06_001]→髄脳(延髄)は脳幹の最尾部で尾方は第一頚神経の根を境として、脊髄に、吻側は橋に移行する。①外表面:外表面には脊髄の前正中裂、前外側溝、後外側溝および後正中溝につづく溝がみられる。前正中裂と前外側溝との間の隆まりは(延髄)錐体とよばれ、錐体路に相当する。錐体交叉は前正中裂を横切って走る線維として外表面からも認められる。前外側溝と後外側溝との間には背側と腹側に隆起がある。腹側の楕円形の隆起はオリーブで、中にはオリーブ核がある。背側の隆起は灰白結節で、三叉神経脊髄路と脊髄路核に相当する。後中間溝と後外側溝の間には楔状束核と薄束核に一致して、外側には楔状束結節と内側には薄束結節とがみられる。さらに上外方には下小脳脚が存在する。②横断面:脊髄との移行部の高さでは、中心管の背側には後索核(楔状束核と薄束核)がある。これらの核からの線維は内弓状線維となり腹内側に向かい交叉する(これを毛帯交叉とよぶ)。交叉後は錐体の背側に集まり内側毛帯を形成する。一方、腹側では錐体交叉をした線維が背外側の側索に入るのがみられる。オリーブ核の高さでは、一般に延髄背側部には基板および翼板由来の脳神経核が配列されている。内側から外側にかけて体性運動性の舌下神経核、一般内臓遠心性迷走神経背側核(内側核)と唾液核がある。同じく基板由来の特殊内臓遠心性の舌咽、迷走、副神経の疑核は腹外方に位置している。さらに、これらの外側には翼板由来の一般内臓感覚性の迷走神経背側核(外側核)、特殊内臓感覚性の孤束と孤束核があり、一般体性感覚性の三叉神経脊髄路核は延髄の最も外側に位置している。その他、副楔状束核が楔状束核の外側に、介在核が舌下神経核の外側にある。これらの神経核の腹側には網様体とよばれる。ここには縫線核がある。延髄背側部で縫線の両側には内側縦束が通り、その腹側には三角形の内側毛帯がある。腹側部には錐体とその背側から外側にオリーブ核がある。なお第四脳室底の中心灰白質の内側部を背側縦束が通る。延髄も上・下行性伝導路を通過させる。また、延髄には第8~12脳神経の諸核、呼吸中枢、循環中枢などが存在し、これらへの圧力(ヘルニア、頭蓋内圧亢進、などによるもの)は昏睡と死をまねく。延髄とはいうのは脊髄の延長部という意味で名づけられたらしい。このラテン名を最初に使ったのはハイステルLorenz Heister(1740)であるが、橋と脊髄の間の部位に限局して使いだしたのはハレルAlbrecht von Haller(1750)である。延髄はその膨らんだ感じから球とも呼ばれる。
Gracile nucleus(薄束核;後索内側(部)核)Nucleus gracilis; Nucleus partis medialis fasciculi dorsalis; Nucleus clavae
[A17_2_06_002]→薄束核はゴル核ともよばれる楔状束核の内側にある核で、細胞学上3つの領域に分けられる、薄束核のニューロンは、三角形、多角形ないし卵円形の中等大の細胞体をもっており、比較的短い、中等度に分岐した少数の樹状突起が各方向に伸びている。核は円形ないし楕円形の切口を示し、しばしば核膜の深い陥入をみる。粗面小胞体は体運動性の細胞のように斑状のニッスル小体をつくることなく、細胞全体に濔漫性に広がり、神経細糸、微細管と入り混じる。遊離のポリゾームも多い。ゴルジ装置はとくによく発達し、核の周りを取り囲み、太い樹状突起のかなり遠位部にまで達している。軸索は、細胞体ないし、むしろしばしば大きな樹状突起の基部から出て神経網に入り、他の薄束核ニューロンの軸索と共に内側弓状線維を形成して内側毛帯に入るのであるが、神経網にはいると間もなく通常1本、時にそれ以上の側副枝を出して、おのれ自身の細胞、ないし同じ薄束核の他の細胞の樹状突起に、自己抑制系(もしかすると促進形)のシナプスるを形成する。薄束を上行してくる後根線維はしばしば糸球体を形成して樹状突起に終わるが、これより数の少ない軸索細胞体終末の中には、錐体路の側副枝が含まれていることが知られている。
Cuneate nucleus(楔状束核;後索外側(部)核)Nucleus cuneatus
[A17_2_06_003]→楔状束核はブルダッハ核とも呼ばれる。脊髄の後索の3核の中の1つ。閂の高さから下方にかけて延髄の背面近くに位置する。楔状束核は薄束核の外側にある核で細胞構築上、部位的差異を示す。楔状束核の背側部には灌木上の樹状突起をもった円い細胞集団の集団があり、腹側部には三角形、多極性、紡錘形の細胞がみられる。円形細胞群は上肢の遠位部から来る一次求心線維を受け、小さな皮膚受容野に関係すると考えられている。Burdach, Karl Friedrich (1776-1847) ドイツの解剖学者、生理学者、ブルダッハ索(脊髄楔状束)を記述(「Vom Baue u. Leben des Gehirns」, 1819-1826)
Cuneate nucleus(楔状束核)Nucleus cuneatus
[A17_2_06_004]→楔状束核はブルダッハ核とも呼ばれる。脊髄の後索の3核の中の1つ。閂の高さから下方にかけて延髄の背面近くに位置する。楔状束核は薄束核の外側にある核で細胞構築上、部位的差異を示す。楔状束核の背側部には灌木上の樹状突起をもった円い細胞集団の集団があり、腹側部には三角形、多極性、紡錘形の細胞がみられる。円形細胞群は上肢の遠位部から来る一次求心線維を受け、小さな皮膚受容野に関係すると考えられている。Burdach, Karl Friedrich (1776-1847) ドイツの解剖学者、生理学者、ブルダッハ索(脊髄楔状束)を記述(「Vom Baue u. Leben des Gehirns」, 1819-1826)
Lateral reticular nucleus(側索核)Nucleus reticularis lateralis; Nucleus funiculi lateralis
[A17_2_06_006]→外側網様体核、外側核ともよぶ。延髄網様体に属する核である。この神経核はオリーブ核群の尾方で始まり、オリーブ中央に分けられる。すなわち、大細胞部、小細胞部、および三叉神経下部である。大細胞部は下オリーブ核の背側方にあるが、小細胞部は網様体格の背外側部にある。外側毛様体核は小脳への中継核であり、脊髄から求心性線維(すなわち脊髄網様体路、および脊髄視床路の側枝)と赤核からの下行線維(赤核延髄路)を受けている。脊髄求心性神経は体部位局在性配列をもって外側毛様体核へ投射している。外側毛様体核の全亜核からきた線維は下小脳脚を通って小脳前葉、半球内側部および小脳核に投射する。その主要な求心性線維は脊髄から両側性にくる両側性腹側屈筋反射路(Bilateral ventral flexor reflex tract)である。その他、大脳皮質運動野などからも線維を受ける。
Medial accessory olivary nucleus(弓状核)Nucleus olivaris accessorius medialis
[A17_2_06_007]→内側副オリーブ核は内側毛帯の外側縁に沿って存在する下オリーブ核の副核。
Olivary nucleus(オリーブ核)Nucleus olivaris おりーぶかく
[A17_2_06_008]→
Medial accessory olivary nucleus(内側副オリーブ核)Nucleus olivaris accessorius medialis
[A17_2_06_009]→内側副オリーブ核は内側毛帯の外側縁に沿って存在する下オリーブ核の副核。
Posterior accessory olivary nucleus; Dorsal accessory olivary nucleus(背側副オリーブ核)Nucleus olivaris accessorius posterior; Nucleus olivaris accessorius dorsalis はいふくおりーぶおりーぶかく
[A17_2_06_010]→背側副オリーブ核は主オリーブ核の背側にある下オリーブ核の副核。
Central gray substance(中心灰白質)Substantia grisea centralis; Stratum griseum centralehypothalami ちゅうしんかいはくしつ
[A17_2_06_011]→中心灰白質は一般用語としては、脊髄の中心管と脳幹の第3脳室、第4脳室に隣接する、またはそれらを取り巻く、主に小形の細胞からなる灰白質をいう。特殊用語としては中脳水道を取り巻く灰白質の厚い袖で、吻方で視床下部の後核に連続しているもをいう。
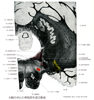
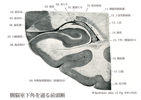
Medullary reticular formation(延髄網様体)Formatio reticularis; Substantia reticularis えんずいもうようたい
[A17_2_06_012]→延髄網様体は系統発生的に脳幹の中で最も古い部分で、脳幹の中心の基質を形成している。菱脳蓋中、迷走神経核、前庭神経核および顔面神経核近くに位置する核。鰓弓および身体の筋肉の調節作用をおこなう。網様体は上方および下方につづいている。
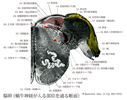

White reticular formation(白網様体)Formatio reticularis alba
[A17_2_06_012_1]→
White reticular substance(白網様質)Substantia reticularis alba はくもうようしつ
[A17_2_06_013]→
Gray reticular substance(灰白網様質;灰白網様体)Substantia reticularis grisea; Formatio reticularis grisea [A17_2_06_014]→
Motor nucleus of facial nerve(三叉神経脊髄路核)Nucleus nervi facialis
[A17_2_06_015]→第7脳神経、すなわち顔面[中間]神経を形成する神経線維のうち、表情筋・広頚筋・アブミ骨筋・茎突舌骨筋などの横紋筋を支配する運動神経線維の起始核であり、橋の最尾側レベルにおいて橋被蓋の腹外側部に位置する。顔面神経核からおこる神経線維は核の背側から出て背内頭側に走り(顔面神経上行根)、第四脳室底の直下で外転神経核の内側部に達してはじめて密な神経束を形成する。ついで、この線維束は外転神経核の頭側レベルで核の背側を外側に向かい顔面神経膝を形成する。ついで、線維束は三叉神経脊髄路核の内側縁に沿うように腹外側に走り(顔面神経下行根)、橋の尾側レベルで脳幹を出る。顔面神経の支配を受ける横紋筋のうち顎二腹筋後腹は副顔面神経核に支配される。副顔面神経核の神経細胞は顔面神経核と三叉神経運動核を結ぶ線上に散在性に存在する。顔面神経に含まれる副交感神経線維の起始核として、上唾液核が記載されている。この核の神経細胞は、橋被蓋網様体の尾側レベルでその背外側部において、三叉神経脊髄路核の内側縁付近に比較的散在性に存在するようである。
Nucleus of glossopharyngeal nerve(舌咽神経核)Nucleus nervi glossopharyngei
[A17_2_06_016]→舌咽神経核 舌咽神経(第9脳神経)の起始核、終止核のことで以下の4つの核からなる。①下唾液核:迷走神経背側核のすぐ吻側にある一般内臓性遠心性線維の起始核で、その神経細胞の軸索は、副交感神経節前線維として耳神経節で節後線維に中継されて耳下腺の分泌に関与する。②疑核:吻側部の神経細胞の軸索は特殊内臓性遠心性線維として茎突咽頭筋と咽頭収縮筋の上部を支配する。③孤束核:吻側部には舌の後1/3の味覚を伝える特殊内臓性求心性線維が終止している。④舌咽神経背側核:迷走神経背側核の吻外側孤束核の内側に位置する神経核で、一般内臓性求心性線維が終止する。(医学書院医学大辞典:大竹一嘉)
Nucleus of vagus nerve(迷走神経核)Nucleus nervi vagi
[A17_2_06_017]→第四脳室底の迷走神経三角の部分に存在する、遠心性線維の起始核と求心性線維の終止核からなり次の三つの核が区別される。(1)迷走神経背側核:これはさらに内側核と外側核とにわけられる。内側核(背側運動核)は一般内臓遠心性である副交感性線維を出す。外側核は迷走神経支配の領域からの一般内臓求心性線維が終止する。この核はまた孤束核に含められ、孤束核の内側核として扱われることが多い。(2)孤束核:孤束の外側にある大型細胞の集団で喉頭蓋領域からの特殊内臓求心性線維である味覚線維が終わる。(3)疑核:延髄網様体でオリーブ核と三叉神経脊髄路核との間に位置する運動性細胞群である。特殊内臓遠心性線維の起始核で、茎突咽頭筋を除く咽頭筋や軟口蓋の筋などの横紋筋を支配する。疑核の吻側部は茎突咽頭筋支配の舌咽神経の特殊内臓遠心線維を、尾側部は副神経延髄根を出す。なお背側核や疑核の周辺には心臓運動抑制線維を出す細胞がある。耳介、外耳道、鼓膜などからの迷走神経の一般体性求心性線維は三叉神経脊髄路核に終わる。(解剖学辞典:松下松雄)
Nucleus of accessory nerve; Accessory nucleus (XI)(副神経核)Nucleus nervi accessorii
[A17_2_06_018]→副神経核は脊髄の上方6区域(C1-C6)の前角の中央部と外側部を縦に連ねた運動性細胞柱で、ここから副神経がでる。副神経は延髄根(内枝)と脊髄根(外枝)からなる。延髄根は疑核の下端部から起こり迷走神経に合して喉頭(固有)筋や下咽頭収縮筋を支配する。脊髄根は延髄下部から頚髄上部(C5-C6)にかけて存在する副神経脊髄核からおこり僧帽筋と胸鎖乳突筋を支配する。脊髄核は延髄下部では前索の内側部の近くにあるが下方にいくにしたがい外方に移動し、前角外側部に位置するようになる。根は背外方に向かい、側索の背側部を貫いて脊髄を出る。
Nucleus of hypoglossal nerve; Hypoglossal nucleus; Nucleus of CN XII(舌下神経核)Nucleus nervi hypoglossi ぜっかしんけいかく
[A17_2_06_019]→舌下神経核は第12脳神経、すなわち舌下神経の起始核である。脊髄前角の運動神経細胞群の頭側延長部として、延髄において第四脳室底の直下で正中線背側部の両側に存在し、オリーブ核下端部より内耳神経核のレベルにわたる細胞柱(約2cm)を形成する。内舌筋および5個のうち4個の外舌筋を支配している運動神経核。
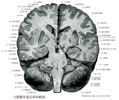
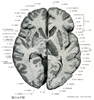
Cerebellum(小脳)Cerebellum
[A17_2_07_001]→Cerebellumは、「大脳、脳」を意味するcerebrumの指小形で、「小さい脳」という意味である。Cerebrumは、「頭」を意味するギリシャ語のkararan由来する。 小脳は筋、関節などの深部組織、前庭、視覚、聴覚系などからの入力を直接あるいは間接的に受け、眼球運動を含む身体の運動調節を司る。小脳は正中部の虫部と外側部の小脳半球とに分けられる。いずれも多数の小脳溝により小脳回に細分される。この中、特定の小脳溝は深く、これにより小脳回の集合ができる。これを小脳小葉とよぶ。ヒトでは小脳は深い水平裂により上面と下面とに分けられ、虫部とそれに対応する半球に九つの小葉が区別される。系統発生的には小脳は前葉、後葉、片葉小節葉の3部分に分けられる。前葉は系統発生的に古く古小脳(Paleocerebellum)ともよばれ、脊髄小脳路、副楔状束核小脳路、オリーブ小脳路の一部、網様体小脳路などをうける。後葉は系統発生的に新しく、新小脳(Neocerebellum)とよばれる。とくに半球部は虫部より新しく、橋核、主オリーブ核などを介して大脳皮質と結合している。前葉と後葉とは第1裂により境される。片葉小節葉は原小脳(Archicerebellum)とよばれ最も古く前庭系との結合が著明である。後葉とは後外側裂で境される。後葉には虫部錐体と虫部垂との間に第2裂がある。ヒトの小脳小葉の形は他の動物のものと大きく異なりる。小脳全体は灰白質と白質とからなる。灰白質には小脳皮質と小脳核とがある。小脳皮質は小脳小葉の表面をなし遠心性軸索を出すPurkinje細胞と皮質内での結合を行う細胞とからなる。小脳核は深部にあり、室頂核、球状核、栓状核、歯状核の4核からなる。小脳皮質にはその結合から三つの縦帯が認められる。すなわち、正中部の虫部皮質、外側部の半球皮質および両者の境界部の虫部傍皮質である。虫部皮質のPurkinje細胞は室頂核に、虫部傍皮質は球状核と栓状核に、半球皮質は歯状核に投射する。小脳の中心部の白質塊は髄体とよばれ、遠心性および求心性繊維から出来ている。ここからは白質が分枝して(白質板)、小葉に分かれる。全体として樹の枝のようにみえるので、小脳活樹と名づけられている。小脳は三つの小脳脚により、延髄、橋、中脳と結合している。これは小脳の遠心路および求心路の通路となっている。 小脳の発生 development of the cerebellum:小脳は後脳の菱脳唇から発生する。後脳の菱脳唇は翼板の背外側につづく背内方に突出する高まりで、胎生2ヶ月の後半において急速に増大し、小脳板とよばれるようになる。左右の小脳板の間には菱脳蓋の頭側半分が介在するので、頭側部では左右の小脳板は相接しているが、尾側部では広く離れている。菱脳の中央部を頂点とする橋弯曲が高度になると、この部の菱脳蓋の左右方向の拡大によって、左右の小脳板の尾側部はいよいよ高度に引き離され、左右の小脳板は菱脳の長軸に直角な一直線をなすようになる。これと同時に左右の小脳板の頭側部(今では内側部)が合一するので、結局、正中部が小さくて左右両部が大きい単一の小脳原基が成立する。正中部からは小脳虫部が、左右両部からは小脳半球が形成される。 増大していく小脳原基の背側部には、やがて中部から半球に向かって走る溝が次々に出現して小脳を区画する。胎生5ヶ月のおわりには小脳虫部における10個の主な区分(小脳葉)がほぼ完成する。これらの小脳葉はそれぞれ固有の発育を行うが、その間に第2次、第3次の溝が生じて、各小脳葉を多数の小脳回に分ける。このような形態発生の結果広大な表面積を獲得した小脳の表面には小脳皮質とよばれる特別な灰白質が形成され、これに出入りする神経線維はその深部に集まって小脳白質を形成する。 小脳原基においても菱脳室に接する内側から表面に向かって胚芽層・外套層・縁帯の3層が分化する。胚芽層は神経が細胞をつくりだすが、胚芽層から発生するのは小脳核の神経細胞と小脳皮質のPurkinje細胞およびGolgi細胞である。小脳原基が3層に分化するとまもなく、外套層の表層部にやや大型の神経芽細胞が出現し、小脳板の背側面(表面)に平行に1列にならぶ。これがPurkinje細胞の幼若型である。ついで小脳板の尾側端部の胚芽層でさかんな細胞分裂がおこり、ここで生じた未分化細胞は縁帯の表層部を頭方に遊走して、小脳原位の全表面をおおう未分化細胞層を形成する。これを胎生顆粒層という。 胎生顆粒層の細胞は胚芽層における細胞分裂が終わるころから活発な分裂を開始し、神経細胞をつくりだす。この神経細胞は縁帯およびPurkinje細胞の層を貫いて、Purkinje細胞の層の下に達し、ここに新しい細胞層(内顆粒層)をつくる。胎生顆粒層からは、このほかに縁帯の中に散在する籠細胞や小皮質細胞が生ずる。必要な数の神経細胞を送り出すと胎生顆粒層における分裂はやみ、本層は速やかに消失する。一方、Purkinje細胞は縁帯の中に多数の樹状突起を伸長させる。縁帯はPurkinje細胞の樹状突起で満たされて厚くなり、核をあまり多く含まな灰白層となる。このようにして小脳の全表面は、表面から灰白層・Purkinje細胞層・内顆粒層の3層から成る小脳皮質でおおわれることになる。
Globose nucleus; Posterior interpositus nucleus(歯状核)Nucleus interpositus posterior; Nucleus globosus
[A17_2_07_002]→球状核は2~3個の円い細胞集団で、栓状核の内側、室頂核の外側に位置する。ここには大型および小型の多極細胞がある。下等哺乳動物では栓状核と球状核は連続しているように見え、これらを一括して中位核とよぶ。その組成細胞および線維連絡の違いにより、これはヒトの、①栓状核に相当する前中位核と、②球状核に相当する後中位核に区別される。
Emboliform nucleus; Anterior interpositus nucleus(栓状核)Nucleus interpositus anterior; Nucleus emboliformis
[A17_2_07_003]→栓状核は楔形の細胞集団で、歯状核門の知覚に位置する。歯状核とよく似た細胞からなり、しばしば両者を区別することが困難である。小脳皮質中間域のPurkinje細胞の軸索を受ける。この核の細胞の軸索は上小脳脚から小脳を出て行く。 TAではAnterior interpositus nucleus; Emboliform nucleusとなっている。
Globose nucleus; Posterior interpositus nucleus(球状核)Nucleus interpositus posterior; Nucleus globosus
[A17_2_07_004]→球状核は2~3個の円い細胞集団で、栓状核の内側、室頂核の外側に位置する。ここには大型および小型の多極細胞がある。下等哺乳動物では栓状核と球状核は連続しているように見え、これらを一括して中位核とよぶ。その組成細胞および線維連絡の違いにより、これはヒトの、①栓状核に相当する前中位核と、②球状核に相当する後中位核に区別される。
Fastigial nucleus; Nucleus medialis cerebelli(室頂核)Nucleus fastigii; Nucleus medialis cerebelli
[A17_2_07_005]→室頂核は小脳核のうち最も内側で、第四脳室上壁の正中線知覚に位置する。核の中で細胞に差があり、小型細胞が腹側を示す。この核の外側縁の細胞は腹外側に伸びて前庭神経核に向かっているが、GolgiⅡ型細胞は存在しないようである。他の小脳核の細胞と異なり、室頂核細胞からは交叉性および非交叉性軸索が出、そのうち交叉性のものは核の吻側部から多く出る。小脳核の細胞はPurkinje細胞とは異なり、促進的で、小脳の外に投射する。免疫組織化学の研究結果から、小脳核の全ての細胞の促進性伝達物質はグルタミン酸塩およびアスパラギン酸塩であるらしいと考えられる。小脳皮質からの唯一の出力であるプルキンエ細胞は一定の配列様式をもって小脳核に投射し、小脳髄質中に存在する細胞群(小脳核)から起こる促進性出力系に対して抑制的に働く。室頂核からの遠心性線維は、①上小脳脚を通らない、②大部分が小脳内で交叉する。③脳幹各部の神経核に投射する、点で特徴的である。室頂核からの交叉性線維は鈎状束を通って小脳の外に出るが、これは上小脳脚の周囲を弓形に走る。また非交叉性線維は傍索状体を通って脳幹に投射する。鈎状束中の交叉性線維は室頂核のすべての部位の細胞から起こり、その数は傍索状体中を走る非交叉性線維より多い。室頂核からの投射線維のうちで最も多いのは下位脳幹へ到るものである。前庭神経核への投射は両側性で、前庭神経外側核および下核の腹側部に対称性に終わる。室頂核網様体線維は主に核の吻側部から起こり、大部分交叉して、①巨大細胞性網様核の内側部、②橋網様体尾側部、③傍正中網様核背側部および④外側網様核の各部に終止する。交叉性の室頂核橋線維は鈎状束から分かれて腹側に走り、橋核の背外側部に終わる。また少数の交叉性室頂核脊髄線維が上部頚髄まで下行し、そこで前柱細胞に接続する。室頂核からの線維のうち、少数のものは脳幹の背外側部を上行し、側枝を上丘および交連核に送り視床の細胞が疎らな部位(VLcとVPLo)に両側性に終わる。これらの終止は歯状核および中位核からの中位核からの終止と重なり合うことはない。
Spinal cord(脊髄)Medulla spinalis おうきゃくそく
[A17_2_08_001]→脊髄は頚部(頚髄)、胸部(胸髄)、腰部(腰髄)、仙骨部または脊髄円錐(仙髄と尾髄)とからなり、それぞれ髄節に分かれ、それに対応して31対の脊髄神経が出る。頚髄では8対の頚神経、胸髄では12対の胸神経、腰仙髄では各々5対の腰神経と仙骨神経とが出る。尾髄からは通常1対の尾骨神経が出る。上肢および下肢支配の神経の出る頚髄下部と腰髄下部は発達が著しく、太くなっており、それぞれ頸膨大、腰部大とよばれる。脊髄下端は細くなり脊髄円錐となっておわる。その高さは成人では第1ないし第2腰椎の高さに相当する。新生児、幼児では低く第3腰椎の高さでおわっている。脊髄円錐の先はさらに細く糸状の終糸となって尾骨の背面に付着している。終糸に沿って走る脊髄神経の束はその形状から馬尾とよばれている。脊髄外側面でその腹側と背側の正中には(前)正中裂および(後)正中溝とよばれる溝があり、脊髄を左右の半分に分けている。前者は後者より深く、そこには前脊髄動脈が走っている。左右の脊髄半の外側面には腹側の前外側溝と背側の後外側溝の二つの溝がある。頚髄の高さの背側面は中心部の灰白質とその周辺の白質から成る。灰白質はそれぞれ前角(柱)、中間質(帯)、後角(柱)がある。灰白質の中央を貫いて中心管が通る。上方は第四脳室に開き、下方は脊髄炎水の所では拡大して終室となる。白質は前外側溝と後外側行と②より腹側の前索と外側の側索および背側の後索の3部分に分けられる。頚髄の高さで後索は後中間溝により内・外の薄束と楔状束とに分けられる。
Grey matter of spinal cord; Grey substance of spinal cord; Gray matter of spinal cord; Gray substance of spinal cord(灰白質)Substantia grisea (Medullae spinalis)
[A17_2_08_002]→脊髄において神経細胞が集団をなして存在する部分で、その横断面の形は高さによって異なるがH型をなしている。灰白質は頭尾方向に柱をなし、さらに背側部の後柱(角)と腹側部の前柱(角)とに分けられる。胸髄と腰髄上では外側部に側柱(角)が認められる。前角と後角の間の部分は中間質(帯)とよばれている。灰白質にある神経細胞の集団は細胞構築学的にRexedが行った第Ⅰ層からⅩ層までの層区分は、機能的全体像がみやすいという点で旧来の、諸柱(または諸核)を個別に命名する方法よりも利用価値が高い。Rexedの各層の番号は、後角尖からはじまって前角に向かう順序となっており、Ⅰ~Ⅹのローマ数字が用いられている。 第1層は後角の帽子に相当するような薄層である。この層のニューロンは後根から脊髄に侵入する温・痛覚線維の一部を受け、反対側の脊髄視床路の成分の一部になるような上行性線維を伸ばす。 第Ⅱ層は旧来名の膠様質に相当する。これのニューロンは痛覚に関係したかなりの量の神経信号を、後根からの線維群ばかりでなく延髄網様体の大縫線核からの下行線維群からも受ける。したがって痛みの調節(セロトニン、ノルエピネフリン、P物質、エンケファリンなど多様な神経伝達物質が使われ、また触覚受容ニューロンも関与する。この層内に存在するニューロン細胞体は、軸索突起を、上行性伝導路に直接伸ばすのではなく第Ⅰ、Ⅳ、Ⅴ層ニューロンにシナプス伝達を行なうために伸ばすだけである。 第Ⅲ層と第Ⅳ層は第Ⅱ層に似てはいるが、しかし第Ⅱ層よりも多数の、後根からの痛覚・温度覚・触覚線維を受ける。また、第Ⅲ、Ⅳ層のなかに第Ⅴ層の大型ニューロン(反対側を上行する脊髄視床路の成分となる軸索突起を伸ばすもの)の樹状突起群が侵入している。上衣の頚髄における第Ⅰ~Ⅳ層は一体化し、三叉神経脊髄路核に移行する。第Ⅴ層は後根からの求心性線維、第Ⅱ・Ⅲ・Ⅳ層の介在ニューロン、などからの神経信号を受ける。第Ⅴ層ニューロンから伸びる軸索突起群は、反対側を上行する温・痛覚と軽微(粗大)な触覚のための伝導路、すなわち脊髄視床路の主要成分をなす。皮質脊髄路、赤核脊髄路からの多くの下行性線維も第Ⅴ層に終わる。 第Ⅵ層はおもに頚膨大と腰仙膨大に存在し、顧客筋からの固有感覚入力を受ける。 第Ⅶ層には、いくつかの大きなⅣ細胞集団が多数の介在ニューロンとともに含まれている。中間質外側核と呼ばれる細胞集団は第1胸髄から第2腰髄までの高さにおける脊髄灰白質の側角をなすものであるが、この細胞集団は交感系節前ニューロン細胞体の第Ⅶ層での集まりにほかならない。中間質内側核という名の細胞集団は脊髄の全長にわたっって第Ⅶ層に存在するものであり、臓性求心性線維群を受ける。胸髄核(クラーク背側核とも呼ばれるもの)は第1胸髄から第3胸髄までの第Ⅶ層にあり、筋紡錘や腱器官などからの固有感覚入力を受ける。胸髄核から発する軸索は同側上行性の後脊髄小脳路を形成する。また、第2~4仙髄での第Ⅶ層には、仙髄副交感核(副交感系の節前ニューロン細胞体の集まり)がある。 第Ⅷ層は筋緊張、姿勢の反射的調節にあずかる前庭脊髄路や網様体脊髄路からの下行線維を受ける。この層のニューロン細胞体から伸びる軸索は、同側および反対側の第Ⅶ層と第Ⅸ層に終わる。 第Ⅹ層では体性遠心性ニューロン(前根から脊髄をでて骨格筋に向かう軸索を有すもの)の脂肪体が内・外側集団をつくる。内側集団(別名:内側核)は体幹筋支配にあずかり、脊髄全長における第Ⅸ層で認められるのに対して、外側集団(別名:外側核)は上・下肢筋支配にあずかる関係で頚膨大と腰仙膨大における第Ⅸ層にしか認められない。アルファ運動性、ガンマ運動性の両方のニューロン細胞体が第Ⅸ層に存在している。 第Ⅹ層は脊髄中心管を囲む領域であり介在ニューロン、神経膠細胞、交叉性軸索がそこに含まれている。
Posterior thoracic nucleus; Dorsal thoracic nucleus(胸髄核;背核)Nucleus thoracicus posterior; Nucleus dorsalis thoracici
[A17_2_08_003]→背核ともよび、一般にClarke柱あるいはClarke背核ともよばれている。T1からL3の高さまで中間帯の背内側に存在する細胞群で大、中、小の細胞から構成されている。大ないし中等大細胞の軸索は同側の側索の背外側部を上行する後脊髄小脳路となる。Clarke柱はその高さの後根線維を受けるが、胸髄上部の高さではC5以下C8の後根線維を、腰髄の高さではL4以下の後根線維を受ける。その入力は主にⅠa群(筋紡錘由来の求心性線維)、Ⅱ群線維(腱器官由来の求心性線維)に由来する。
Lateral intermediate substance; Lateral intermediate grey matter of spinal cord; Lateral intermediate gray matter of spinal cord(中間質外側核;側柱核)Substantia intermedia lateralis (Medullae spinalis)
[A17_2_08_004]→中間質外側核は、胸髄および上部腰髄(T1からL2ないしL3まで)の側角の先端部分にある神経細胞柱である。この核の神経細胞は紡錘形で、交感神経性の節前線維を出している。その交感神経線維は前根より出て白交通枝を経て、いくつもの交感神経節に到達する。
Gelatinous substance(膠様質;後柱膠様質)Substantia gelatinosa こうようしつ
[A17_2_08_005]→後角尖の背側にある明るい部分。難染性の神経膠細胞および少数の小神経細胞よりなる。 (Feneis)
Extrapyramidal system; Extrapyramidal tract(錐体外路系;錐体外路)Systema extrapyramidalis すいたいがいろけい;すいたいがいろ
[A17_3_01_001]→錐体路以外の下行性伝導路の総称。機能的には随意運動を制御する神経系をさし、大脳核・小脳・中脳(赤核・黒質)・網様体などから錐体路に至る神経路を意味する。 (335)錐体路(皮質脊髄路+皮質延髄路)以外の運dの迂路の総称。大まかにいえば「感覚情報に応じて随意運動(錐体路)を制御するシステム」で、次のような系統がある。(イラスト解剖学) 錐体外路 運動に関係がある下行性神経路で、錐体路以外のものをまとめて錐体外路という。 いわゆる随意運動で、とくに敏速巧妙な運動は錐体路によるが、そのほかは錐体外路が関与する。たとえば、歩行はほとんど意識しないのに円滑に行われる。このように意識いで歩行が行われるのは、多くの筋群の緊張・弛緩、興奮・抑制などが調節的に調節さえているからである。このような無意識的な巧みな骨格筋の調節作用を行うのが錐体外路系である。 錐体路系がα運動ニューロンを支配するのに対して、錐体外路系はα運動ニューロンのほかにγ運動ニューロンを支配し、筋の緊張状態を調節すると考えられている。 鳥類以下の動物では、骨格筋は錐体外路系で支配され、哺乳類になると錐体路系が発達して骨格筋の支配に加わる。 錐体外路系の神経路は脊髄で前索・側索を散在性に下行する。 つぎのような下行性神経路がある(図9-18)。 ◇図9-18.脊髄における下行性神経路 a.視蓋脊髄路tractus tectospinalis 中脳(上丘)から起こり、交叉して前索を下行する。主として視覚刺激に対応して行う反射的な姿勢・体位の維持や運動と関係がある。 b.赤核脊髄路tractus rubrospinalis 錐体側索路の前側にあるが、ヒトでは発達が悪い。筋緊張の調節にあずかるといわれる。 c.前庭脊髄路tractus vestibulospinalis 前索を下行する。平衡覚刺激に応じて、前庭器や小脳からのインパルスを脊髄に伝え、反射的に緊張節を行う(姿勢反射)。 d.網様体脊髄路tractus reticulospinalis 橋・延髄の網様体から起こり、前・側索を下行する。運動ニューロンに対して抑制や促進作用をもつ。また、不随意的な呼吸運動の調節などにもあずかるといわれる。(解剖学講義)
Cortical extrapyramidal system(皮質錐体外路系)Systema coricales extrapyramidales system ひしつすいたいがいろけい
[A17_3_01_002]→大脳皮質から脳幹各部に線維を送る経路。他の錐体外路系の調節にあずかる。(イラスト解剖学)
Striopallidal extrapyramidal system(線条体淡蒼球錐体外路系)Systema striopallidares extrapyramidales せんじょうたい-たんそうきゅうすいたいがいろけい
[A17_3_01_003]→大脳核(線条体・淡蒼球)から脳幹各部に線維を送る経路。錐体外路系の腫部をなし、骨格筋の調節に重要な役割を果たす。(イラスト解剖学)
Cerebellar extrapyramidal system(小脳錐体外路系)Systema cerebellares extrapyramidales しょうのうすいたいがいろけい
[A17_3_01_004]→小脳と大脳核や脳幹各部を連絡し、運動のコントロールに働く経路。(イラスト解剖学)
Mesencephalospinal extrapyramidal system(脳幹脊髄錐体外路系;中脳脊髄錐体外路系)Systema mesencephalospinales extrapyramidales のうかん-せきずいすいたいがいろけい
[A17_3_01_005]→中脳から脊髄に線維を送り、上記三系統と脊髄運動ニューロンの間を連絡する経路。これにより、脊髄前角の運動ニューロンは、直接あるいは間接的にコントロールされる。(イラスト解剖学)
Peripheral extrapyramidal system(末梢錐体外路系)Systema peripheres extrapyramidales
[A17_3_01_006]→

You are the th visitor to this page !(since 2000/1/1)
本日の番目です。昨日は
th visitor to this page !