(中枢神経系の伝導路)Tractus systematis nervorum ventralis
[A17_0_00_000]→中枢神経系において、末梢から中枢への情報や中枢から末梢への指令の伝達はニューロンの連鎖によってつくられる。上行性伝導路と下行性伝導路に大別される。
Association pathway; Association tract(連合神経路;聯合神経路)Tractus nervosi associationis れんごうほうせんろ [A17_1_01_001]→
Arcuate fibres; Arcuate fibres of cerebrum(大脳弓状線維;弓状線維)association pathway
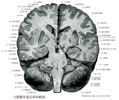
[A17_1_01_002]→弓状線維(束)は島の周囲をこれに沿って走り、終末は扇状に放散して上、中前頭回と側頭葉の一部とを結合する。この線維束中の上方に位置している線維群は後方へ伸びて頭頂葉と後頭葉に達し、上縦束と呼ばれる。
Superior longitudinal fasciculus; Arcuate fasciculus(上縦束;前頭後頭束)Fasciculus longitudinalis superior; Fasciculus arcuatus; Fasciculus frontooccipitalis
[A17_1_01_003]→上縦束は連合線維の中で最長で大脳半球の半卵円中心の外側部にある。頭葉、後頭葉、および側頭葉を結んでいる。下および中前頭回から起こって島の上端の近くを走り、一部は後走して後頭葉内に放散し、一部は弓状に曲がって被殻の周囲に向かい、側頭葉の前端へと走る。
Inferior longitudinal fasciculus(下縦束;側頭後頭束)Fasciculus longitudinalis inferior; Fasciculus temporooccipitalis
[A17_1_01_004]→下縦束は側頭後頭束ともいわれ、側頭葉の前部から起こり、側脳室の下角および後角の外側をはしり、後頭葉の後端部に至る。これは後部では外矢状層を通る。外矢状層にはなお視床から後頭葉に至る視放線の線維が含まれる。また外矢状層と壁板との間には内矢状層があり、これは主として後頭葉から上丘、外側膝状体などに至る線維からなる。
Uncinate fasciculus(鈎状束)Fasciculus uncinatus cerebri
[A17_1_01_005]→鈎状束は島限の下にある緻密な線維束で前頭葉の眼窩回と側頭葉の前部とを結ぶ。この線維束の深部は前頭葉と後頭葉とを連絡する(下後頭前頭束という)。
Cingulum(帯状束)Cingulum prosencephali
[A17_1_01_006]→帯状束は帯状回の白質内に縦に通っているよく目立つ半球内側面の主要な連合線維束。前頭葉および頭頂葉の内側領域と海馬傍回および隣接の側頭葉皮質領域とを連結する、種々の長さの線維を含んでいる。
Fasciculi proprii of spinal cord(脊髄固有束;節間束)Fasciculi proprii こゆうそく(せきずいの)
[A17_1_01_007]→
(固有外側束)Fasciculus lateralis proprius
[A17_1_01_008]→
Commissural pathway(交連神経路)Tractus nervosi commissurales こうれんしんけいろ
[A17_1_02_001]→
Corpus callosum(脳梁)Corpus callosum
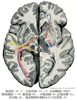
[A17_1_02_002]→脳梁は左右の大脳皮質、ことに新皮質を結合する線維の集合したもので、系統発生的には最も新しく、ヒトでは非常に発育がよい。その前後経はほぼ7.7cmである。脳梁は正中断では全体としては釣針状で、4つの部分が区別される。後端部は膨大し、脳梁膨大といい、その前方に続いて水平に走る部分を脳梁幹とよぶ脳梁はその前端では強く屈曲し、脳梁膝をつくる。これはさらに後下方にくちばしのように尖って脳梁吻となり、しだいに薄くなって終板に続く。
Radiation of corpus callosum(脳梁放線)Radiatio corporis callosi
[A17_1_02_003]→脳梁放線は左右の大脳半球の半卵円中心に放射する脳梁の線維群。脳梁の線維群は大脳縦裂の床に当たる部位を横断し、側脳室の天井の大部分を形成しており、種々の皮質領域に分布する時、豊富な脳梁放線として扇状に広がる。
Frontal part of coronary radiation(前頭部(脳梁放線の))Pars frontalis (Radiatio corporis callosi) ぜんとうぶ(のうりょうほうせんの)
[A17_1_02_003_1]→
Parietal part of coronary radiation(頭頂部(脳梁放線の))Pars parietalis (Radiatio corporis callosi) とうちょうぶ(のうりょうほうせんの)
[A17_1_02_003_2]→
Temporal part of coronary radiation(側頭部(脳梁放線の))Pars temporalis (Radiatio corporis callosi) そくとうぶ(のうりょうほうせんの)
[A17_1_02_003_3]→
Occipital part of coronary radiation(後頭部(脳梁放線の))Pars occipitalis (Radiatio corporis callosi) こうとうぶ(のうりょうほうせんの)
[A17_1_02_003_4]→
Minor forceps; Frontal forceps; Anterior forceps(小鉗子)Forceps minor; Forceps frontalis
[A17_1_02_004]→小鉗子は脳梁放線線維の前方部分でU字形をして脳梁膝を通り、左右の後部前頭葉の連絡をおこなう。
Major forceps; Occipital forceps(大鉗子)Forceps major; Forceps occipitalis
[A17_1_02_005]→大鉗子は脳梁放線線維の後方部分で、脳梁膨大の中をU字形に走る線維。後部後頭葉の連絡をおこなう。
Anterior commissure(前交連)Commissura anterior
[A17_1_02_006]→間脳の前交連は第三脳室の前壁をつくる終板の後ろにある横走線維束である。前部は小線維束で、左右の両側の嗅脳系を結び、後部は大きな線維束で、左右両側の側頭葉に連絡する。前交連は小さな密な線維束で、脳弓柱の吻側で正中線を横切る。これは全体として自転車のハンドルに似た形態をしていて、肉眼標本では明瞭ではないが、2つの部分から構成される。前交連の小さい前部は肉眼標本ではあきらかでないが両側の嗅球を連絡している。大きい後部は主として両側の中側頭回および下側頭回の間を連絡する。
Anterior part of anterior commissure(前部)Pars anterior commissurae anterioris
[A17_1_02_007]→前交連の前部は小線維束で、左右の両側の嗅脳系を結ぶ。
Posterior part of anterior commissure(後部)Pars posterior commissurae anterioris
[A17_1_02_008]→前交連の後部は大きな線維束で、左右両側の側頭葉に連絡する。
Posterior commissure(後交連)Commissura posterior; Commissura epithalamica
[A17_1_02_009]→後交連は、中脳と間脳の背側における境界をなす。この小さな交連は中脳水道と第三脳室の移行部で上丘の上方の中心灰白質の背側にある。交連線維は外側で扇状にひろがり、その周囲を後交連核と総称される細胞が取り囲む。これまで明らかにされた後交連の構成線維は、①視蓋前核、②後交連核、③間質核からの線維がある。瞳孔の対光反射に関与する線維は後交連で交叉する。後交連の下方の中脳水道上衣は線毛をもつ高い柱状細胞よりなる。このような変形上衣細胞は交連下器官を形成し、分泌機能をもち、脳室周囲器官の一つとされている。交連下器官は中脳において脳血液関門をもたない唯一の部分である。
Habenular commissure(手綱交連;手綱交連;松果交連)Commissura habenularum; Commissura pinealis
[A17_1_02_010]→手綱交連は左右の手綱核からの線維の交叉。交叉部は松果陥凹の上方にある。
Commissure of fornix(脳弓交連)Commissura fornicis
[A17_1_02_011]→脳弓交連は海馬交連ともよばれる。脳梁後部の下で脳弓脚間にできる三角形の結合野。多数の線維が反対側の脳弓脚へ行き海馬に終わっている。人では発達がわるい。
Supraoptic commissures(視交叉上交連)Commissurae supraopticae
[A17_1_02_012]→
Dorsal supra-optic commissure(背側視交叉上交連)Commissura supraoptica dorsalis
[A17_1_02_013]→背側視交叉上交連は、さらに背側部(Ganser交連)と腹側部(Meynert交連)に分けられる。腹側視交叉上交連はGudden交連に相当する。これらは交連ではなくて交叉であると思われる。ヒトではその線維の由来及び終止は明瞭でない(昭和60)。
Ventral supra-optic commissure(腹側視交叉上交連;グッデン交連)Commissura supraoptica ventralis
[A17_1_02_014]→Gudden交連ともよばれている。腹側視交叉上交連は一部は視交叉中にある。両側の内側膝状体の連絡らしい。グッデン Gudden, Bernhard Aloys von (1824-1886)ドイツの神経学者、精神科学者。ミュンヘン大学教授樹。Gabersee精神病院長であったが、ここでルードィッヒ3世の治療中、王の入数自殺に巻き込まれて死ぬ。グッデン交連(下交連)グッデン神経節(Ggl. Interpedunculare)を記述。
Tegmental ducussations(被蓋交叉)Decussationes tegmentales
[A17_1_02_015]→
Projection pathways(投射神経路)Tractus nervosi projectionis とうしゃしんけいろ
[A17_1_03_001]→
Corticothalamic fibres(皮質視床路)Fibrae corticothalamici
[A17_1_03_002]→皮質視床線維は大脳皮質のいたる所から出て視床に終わる線維の総称。
Thalamocortical fascicle(視床皮質路;視床皮質束)Tractus thalamocorticalis ししょうひしつろ;ししょうひしつそく
[A17_1_03_003]→
Central tegmental tract(中心被蓋路;視床オリーブ路)Tractus tegmentalis centralis
[A17_1_03_004]→中心被蓋路は赤核尾端からオリーブ核頭端にかけて網様体のほぼ中央部を縦走する線維束である。大部分の線維は小細胞性赤核におこり、同側の主オリーブ核におわる、とされている。線維束をその形状や位置で命名する場合には一般にfasciculusを用い、起始と終止で命名する場合にはtractusを用いることが多い。Tractus rubroolivarisはFasciculus tegmentalis centralisの主要な構成要素であるが、おそらく上行性の線維も含まれていると考えられる。小細胞性赤核を破壊してナウタ法でみると、大細胞性網様体にも終止性変性線維が認められる。これを赤核網様体路と呼ぶこともある。
Corticohypothalamic tract; Corticohypothalamic fibers(皮質視床下路)Tractus corticohypothalamici; Fibrae corticohypophalamicae
[A17_1_03_005]→
Corticomamillary tract(皮質乳頭路;皮質乳頭体路)Tractus corticomamillaris ひしつにゅうとうろ;ひしつにゅうとうたいろ
[A17_1_03_006]→
Corticohabenular tract; Corticohabenular projections(皮質手綱核路;皮質手綱路)Tractus corticohabenularis ひしつたづなろ
[A17_1_03_007]→
Corticotegmental tract; Corticotegmental fibres(皮質被蓋路;皮質被蓋線維)Tractus corticotegmentales; Fibrae corticotegmentales ひしつひがいせんい
[A17_1_03_008]→脳の高位レベルから起こる神経線維の中にも脳幹網様体外側部のニューロンに終止するものがある。これらのなかで重要なものは反対側の台の皮質運動領から起こる皮質被蓋線維と、反対側の赤核から起こる神経線維である。
Corticopontine fibres; Corticopontine tract(皮質橋核路;皮質橋路;皮質橋線維)Tractus corticopontinus
[A17_1_03_009]→皮質橋核路は錐体路と併走しつつ下行し、しだいに橋核に終わる。これに接続する橋小脳路は大部分交叉して橋を横走し(横橋線維)、主として反対側の中小脳脚を通って小脳皮質に終わる。とくに橋核の内側ないし背内側部は橋虫部へ、外側部は小脳半球へ投射する。
Frontopontine fibres(前頭橋(核)路)Tractus frontopontinus
[A17_1_03_010]→前頭橋線維は前頭葉、特に中心後回より起こる皮質橋伝導路。大脳脚の内側1/6のところにあり、前頭葉と橋を結ぶ。
Parietopontine fibres; Parietopontine tract(頭頂橋(核)路)Fibrae parietopontinae; Tructus parietopontinus
[A17_1_03_011]→頭頂、後頭および側頭橋線維で、頭頂葉、後頭葉および側頭葉をなどから出て内方を通って下行し、大脳脚をへて同側の橋核終わる。
Temporopontine fibres; Temporopontine tract(側頭橋(核)路)Fibrae temporopontinae; Tractus temporopontinus
[A17_1_03_012]→頭頂、後頭および側頭橋線維で、頭頂葉、後頭葉および側頭葉をなどから出て内方を通って下行し、大脳脚をへて同側の橋核終わる。
Occipitopontine fibres; Occipitopontne tract(後頭橋(核)路)Fibrae occipitopontinae; Tructus occipitopontinus
[A17_1_03_013]→頭頂、後頭および側頭橋線維で、頭頂葉、後頭葉および側頭葉をなどから出て内方を通って下行し、大脳脚をへて同側の橋核終わる。
Pontocerebellar fibers(橋核小脳路)Tractus pontocerebellares [A17_1_03_014]→
Transverse pontine fibres(横橋線維)Fibrae pontis transversae
[A17_1_03_015]→横橋線維は大脳橋小脳路の横行線維。皮質橋路→橋核→橋小脳路という経路のうち、橋内での横走線維の名称である。
Superficial pontine fibres(浅橋線維)Fibrae pontis superficiales せんきょうせんい[A17_1_03_016]→

Deep pontine fibres(深橋線維)Fibrae pontis profundae しんきょうせんい[A17_1_03_017]→
Corticonuclear fibres(皮質核路;皮質核線維;皮質延髄路)Fibrae corticonucleares; Tractus corticonucleares
[A17_1_03_018]→皮質核線維(皮質延髄路)は大脳皮質の一次運動野(中心前回の下1/3部)から起こり、内方に向かって集まり、その膝を通って大脳脚に入り、その内側部を下行する。線維は脳神経の運動核(動眼神経核・滑車神経核・外転神経核・三叉神経運動核・顔面神経核・疑核・舌下神経核)に終わる。線維は大部分反対側に交叉するが、一部は非交叉性で同側の運動核終わる。直接に運動核に終わる線維のほかに、毛様体にある介在ニューロンを経て運動核のニューロンに接続するものも多い。運動核の運動ニューロンは、それぞれ脳神経として頭頚部の筋(眼筋・表情筋・咀嚼筋・咽頭後頭の筋・舌筋)を支配する。
Corticotectal tract(皮質視蓋路)Tractus corticotectalis [A17_1_03_019]→
Corticospinal fibres: Corticospinal tract(皮質脊髄路;錐体路)Fibrae corticospinales; Tractus corticospinalis
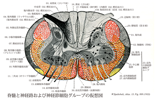
[A17_1_03_020]→①皮質脊髄線維(皮質脊髄路)は大脳皮質の一次運動野(中心前回の中央部と上部)から起こり、内包に向かって集まり、内包の後脚を通って下行する。内包では、上肢に対する線維は線維は後脚の前部を、下肢に対する線維は後部を走る。ついで、大脳脚に入り、その中央2/3部を下行し、橋・延髄に至る。延髄では、その下部の腹側中央部にあつまり、錐体(実際はその一部)を形成する。それで皮質脊髄線維は錐体路ともいわれる。延髄の下端(大後頭孔のすぐ上方)で、線維は反対側に交叉して錐体交叉をつくる。錐体をつくる下行性線維の大部分は錐体交叉で反対側に交叉し、脊髄側索を外側皮質脊髄路(錐体前索路)として下行する。交叉する線維の割合は個人差が大きい。また、約75%の人で交叉する割合が左右非対称で、左側の錐体路の方が交叉する割合が大きい。外側皮質脊髄路の線維は脊髄を下行しつつ、脊髄灰白質に入り、前角の運動ニューロンに接属する。前角の運動ニューロンに直接に終わる線維と、介在ニューロンを経て関節に連絡するものとがある。前皮質脊髄路の線維は脊髄を下行し、前交連を通って交叉し、反対側に終わるが、一部は非交叉性で同側に終わる。このように皮質脊髄路は大脳皮質からおこり、脊髄前角に達し、その運動ニューロンへ運動指令を伝え、骨格筋の運動を起こさせる。延髄の錐体を通る伝導路のうちで、中心前回の一次運動野から起こる線維は約40%で、頭頂葉とくに中心後回や傍中心小葉などから起こる線維が約30%、前頭葉の運動前野などから発する線維が約30%を占めるといわれる。頭頂葉から発する線維は後索核や脊髄後角の膠様質などに達し、知覚性インパルスの流入に対して調整的な働きをするともいわれる。 ②皮質脊髄路は皮質から脊髄への線維束複合で脊髄内を下行し外側皮質脊髄路および全皮質脊髄路を形成する線維は大脳皮質第5層の錐体細胞から発したもので、中心前運動野(Brodmannの4野)、運動前野(6野)と少数ながら中心後回からも出ている。4野の起始細胞はBetzの巨大錐体細胞である。ここからの繊維は内包を下行し大脳脚の中央1/3を通り橋腹側部を経て脊髄腹側に錐体として現れる。さらに下行する際大部分の繊維は錐体交叉で反対側に移り脊髄側索の背側半を外側皮質脊髄路となって下行し脊髄全長の灰白質中間帯の介在ニューロンに分布する。体肢に関係する脊髄膨大部では特に手や手指、足や足指の運動に関わる体肢筋を支配する運動ニューロンに直接連絡している。錐体交叉で反対側に移らない少数の繊維は前皮質脊髄路となって同側の脊髄前索を下行し前角の内側半の介在ニューロンに終止する。皮質脊髄路線維がその起始皮質またはその下方で障害されると反対側の体運動に支障を生じ、特に腕や足で深刻となり筋力低下、痙攣が起こる。Babinskiの徴候このような片麻痺の状態に起因する
Decussation of pyramids; Motor decussation of pyramids(錐体交叉)Decussatio pyramidum
[A17_1_03_021]→錐体交叉は延髄の端部つまり脊髄延髄移行部に大きな特徴として見られる。この神経線維は中心灰白質の前方で大きい神経束を作って交叉し、前角の基部を通って後外方へ投射する。互いに入り組んだ神経線維は、時には非常に太く、後下方に向かって走るので横断切片では大部分の神経交叉が斜めに切断される。外側皮質脊髄路の交叉線維は側索の後部を下行し、一方前皮質脊髄路の非交叉性線維は前索を下行する。上方に向かって連続的に見ると皮質脊髄路の交叉は逆の順序で認められる。身体半側の随意運動は反対側の大脳皮質からくるインパルスによって支配される。その解剖学的な証拠として、錐体交叉がある。
Anterior corticospinal tract; Ventral corticospinal tract; Anterior pyramidal tract(前皮質脊髄路;錐体前索路)Tractus corticospinalis anterior; Tractus pyramidalis anterior
[A17_1_03_022]→錐体路線維は大脳皮質から起こり、内包の後脚を通り、次いで大脳脚ではその中央2/3の部分を占める。ここでは錐体路は皮質橋核線維を伴っている。錐体路線維は橋では多数の線維群に分かれ、横走する橋核小脳路として交叉して走る。延髄では、これらの線維は再び一つにまとまり、錐体を形成する。延髄と脊髄の移行部で約80%の錐体路線維が交叉する(錐体路交叉)。その結果、交叉性の外側脊髄路が脊髄の側索を、非交叉性の前皮質脊髄路が前索を通って脊髄へ下行する。注意すべき点は、皮質脊髄路線維は内包の後脚を通るさい、皮質皮質下脊髄路線維や視床大脳皮質線維など、多くの線維と一緒に走るということである。したがって、しばしば起こる内包の血管損傷のさいにみられる症状は皮質脊髄路線維の遮断のみその原因があるわけではない。
Ventrolateral pyramidal fasciculus; Anterolateral pyramidal fasciculus(前外側錐体束)Fasciculus pyramidalis ventrolateralis (Barnes) ぜんがいそくすいたいそく
[A17_1_03_022_1]→
Lateral corticospinal tract; Lateral pyramidal tract(外側皮質脊髄路;錐体側索路)Tractus corticospinalis lateralis; Tractus pyramidalis lateralis
[A17_1_03_023]→錐体側索路 錐体路線維のうち、錐体交叉で交叉する線維で構成される神経路をいう。反対側の脊髄側索後部(後側索ないし背側索)を下行し、途中で漸次脊髄灰白質に線維を出しながら、次第に小さな線維束となり脊髄下端まで達する。通常、9割以上の錐体路線維が交叉性で錐体側索を通ると考えられているが、これら交叉性の錐体路線維と錐体前索路を通る非交叉性のものとの割合は、特にヒトでは個体差が著しい。[医学大辞典:高田昌彦] 外側皮質脊髄路は脊髄全長にわたって下行し、全髄節の灰白質に線維を送り、尾方へ行くに従って徐々に小さくなる。下位腰髄と仙髄で後脊髄小脳路の下方では、外側皮質脊髄の線維が脊髄の背外側表面に達する。交叉性外側皮質脊髄路音線維は中間部で脊髄灰白質に入り、第Ⅳ、Ⅴ、Ⅵ、Ⅶ層の部分に分布する。サルでは少数の線維が直接前角細胞あるいは第Ⅸ層内のその突起に終わる。
(線条体放線)Radiatio corporis striati せんじょうたいほうせん [A17_1_03_024]→
(皮質線条部)Pars corticostrialis ひしつせんじょうぶ [A17_1_03_025]→
(線条視床部)Pars striothalamica せんじょうししょうぶ [A17_1_03_026]→
(線条視床下部)Pars striohypothalamica せんじょうししょうかぶ [A17_1_03_027]→
Optic tract(視索)Tractus opticus
[A17_1_03_028]→視索は視交叉と外側膝状体の間の視覚路で左右の視索は視床下部と大脳脚基底部を回って後外方へ走る。これらの線維の多くは外側膝状体の中に終止するが、小部分は下丘腕となって上丘および視蓋前域にまで続く。外側膝状体からは膝状体鳥距路が起こり、これが視覚路の最後の中継路をなす。視索前域は対光反射と関係し、上丘は眼と頭の反射運動より視覚刺激を追跡することと関係している。網膜視床下部線維は、両側性に視床下部の視神経交叉上核に終止する。この網膜からの直接の投射は、機能的には神経内分泌調節と関連している。
(視索放線)Radiatio tractus optici しさくほうせん [A17_1_03_029]→
(中脳部(視索放線の))Pars mesencephalica radiatio tractus optici ちゅうのうぶ(しさくほうせんの) [A17_1_03_030]→
(膝状体部(視索放線の))Pars geniculata radiatio tractus optici しつじょうたいぶ(しさくほうせんの) [A17_1_03_031]
(視床部(視索放線の))Pars thalamica radiatio tractus optici ししょうぶ(しさくほうせんの) [A17_1_03_032]→
Optic radiation(視放線)Radiatio optica
[A17_1_03_033]→外側膝状体鳥距線維は外側膝状体の小細胞性部の細胞から起こり、内包のレンズ後部を通って視放線を形成する。これらの線維は鳥距溝の上、下両側で、有線野の皮質に終枝する。視放線のすべての線維が最短経路を通って皮質に達するわけではない。最も背方にある線維はほとんどまっすぐに後方に進み有線野に達するが、より腹方にある線維はまず前方、かつ下方に向きを変えて側頭葉の中に入って、側脳室の下角の吻側部の上に拡がり次いで後方に曲がり、側脳室の外壁に接して走り(外矢状層)、後頭葉の皮質に達する。最も腹方の線維が最も長い弧状の経路をとる。網膜の各領域は外側膝状体と正確な点対点の対応関係をもち、網膜の各部分は外側膝状体の局所的に限局された特殊な部分に投射する。網膜の上内側および上外側の両1/4の領域(視野の下半分に該当する)からの線維は外側膝状体の内側半に、また下内側および下外側の両1/4の領域からの線維は外側膝状体の外側半に終止する。黄斑からは、水平経線を通る面の両側で、外側膝状体尾尾側部において、クサビ形を領域に投射する。単眼視による三日月形の視野も含めて、周辺視野からの投射は、外側膝状体の吻側方に終わり、水平経緯線を越えて連続している。同様な点対点の関係は外側膝状体と有線野の皮質の間にも存在する。上外束の両1/4の領域(視野の下半分)に対応する外側膝状体の内側半は鳥距溝の上唇に投射し、しかも、これらの線維は視放線の下部を占める。黄斑からの線維は視放線の中間部を構成し、鳥距溝を囲む大脳皮質の尾方1/3の領域に終止する。網膜の中心傍野および周辺部からの線維はそれぞれ、更に吻側方の部位に終わる。網膜の一定のの場所に光点を停止させた実験的研究によって、次のことが明らかにされた。すなわち、網膜における視神経節細胞の受容野は同心円上に配列している。網膜においては、受容野は受容器である杆状体と錐状体ならびに単一神経節細胞の興奮性に影響を与える網膜の他のニューロンと関係している。網膜は、そこに存在する神経節細胞の数と同じほどの多くの受容野はいずれも次の2つの領域に分けられる。すなわち、①円形の小さい中心領域および②それを取り囲む同心性の領域で、周辺部、あるいは包囲部とよばれる部分である。これら2つの領域は機能的には拮抗している。受容野には一般に次の2型が記載されている。すなわち、①“開”を中心に“閉”を周辺に有する受容野および②“閉”領域を照射すると、神経細胞は活発に発火することになろう。もし、光刺激が、互いに抑制作用を示す“開”および“閉”の両領域を照射すると、それらの刺激は互いに無効になる。網膜の連絡は神経節細胞のレベルにおける同心円上受容野によく一致する。中心部からのインパルスは受容細胞、双極細胞および神経節細胞の間の直接結合によって伝達されるといわれ、他方、おれと拮抗的に働く周辺の領域は、無軸索細胞によって仲介される結合(すなわち、双極細胞と神経細胞の間の結合)を有する。外側膝状体のニューロンの受容野は光りの点を網膜の一定の場所に停止させた場所と類似しているように思われる。しかし、外側膝状体のニューロンは周辺の受容野に比較的大きな抑制を示す。外側膝状体の各層にある細胞は、同側、あるいは反対側、いずれかの一眼の受容野からの投射を受ける。外側膝状体の中の細胞で、両眼からの影響を受けるものは、たとえあるとしても、きわめて少ない。外側膝状体と有線野の細胞との間には受容野固有の重要な変形が生じる。有線野の細胞が“裂隙”光、あるいは、特定の方向に働く視覚模様に敏感である。有線野の中で起こる変形は、いろいろな作動様式をもった皮質細胞の柱状配列によるものであり、これらのはたらきによって、垂直形と水平形の中でのいろいろな変化が記号化される。
(視床枕皮質路)Tractus pulvinocorticalis radiatio optica ししょうちんひしつろ(しほうせんの) [A17_1_03_034]→
Geniculocorticalis tract(膝状体皮質路(視放線の))Tractus geniculocoricalis radiatio optica しつじょうたいひしつろ(しほうせんの) [A17_1_03_035]→
Acoustic radiation(聴放線;膝状体皮質路)Radiatio acustica; Tractus geniculocorticalis しつじょうたいひしつろ(ちょうかくの)
[A17_1_03_036]→聴胞線の線維は内側膝状体から内方の後脚の下部を横切って走り、側頭葉の中を垂直に上って行き聴覚皮質に達する。これらの線維はその際ある局在的配列を守っており、それに伴って内側膝状体の個々の部分は聴覚領の特定の領域へ投射している。聴放線聴放線の経過中にラセンを描く回旋が起こるために、内側膝状体の吻側部は聴覚領の尾側部へ、内側膝状体の尾側部は聴覚領の吻側部へ投射することになる。この回旋はサルでは実験的に、ヒトでは髄鞘形成の経過を追うことによって証明されている。
Transverse crural tract(横脚束)Tractus cruralis transversus; Tractus peduncularis transversus おうきゃくそく [A17_1_03_037]→
Corona radiata; Coronary radiation(放線冠)Corona radiata
[A17_1_03_038]→放線冠とは、投射線維(上行性線維と下行性線維とからなる)で内包から大脳皮質(あるいは大脳皮質から内包)までの神経線維が視床の近くでは投射線維が密で大脳皮質にちかくなるにつれて密度が相対的に粗になるので放散しているように見えることから、放線冠と呼ばれている。
Supra-opticohypophysial tract(視索上核下垂体路)Tractus supraopticohypophysialis
[A17_1_03_039]→視索上核下垂体路は視索上核および室傍核の大型細胞要素で構成されている。これらの大型ニューロンの特異的なペプチド、オキシトシンとバソプレッシンは脳で最初に分離された特徴あるペプチドである。これらのペプチドは常に、ニューロフィジンという大型のペプチドと結合している。ニューロフィジンは分枝前駆体の部分である。ニューロフィジンⅠはオキシトシンに関係し、ニューロフィジンⅡはバソプレッシンと関係がある。2種のニューロフィジンは視索上核と室傍核の中に存在しているが、オキシトシンとバソプレッシンとは異なるニューロンに含まれる。神経分泌物は細胞体内で合成され、軸索内を終末まで輸送される。神経分泌物質は直径120~200nmの果粒が集まってできている。オキシトシンとバソプレッシンとはそれぞれ異なる側面像をもつ軸索内に存在している。視床下部下垂体路のホルモンは終末の軸索の膨隆部の中に、内部の密な有芯小包として蓄積される。密な有芯小包の内容物質は有窓毛細血管が含まれる血管周囲腔の中に放出される。室傍核の大細胞性要素からの第二次遠心性投射線維は正中隆起の外部帯までは説明されいる。そのうえ、高いレベルのバソプレッシンが下垂体門脈系に証明されている。正中隆起の外部帯のバソプレッシンの役割はわかっていないが、多分副腎皮質刺激ホルモン(ACTH)放出の調節に働くのであろう。視索上核と室傍核の細胞はまた、エンケファリン、コレシストキニン、グルカゴン、ディノルフィンおよびアンギオテンシンのような他の生物学的に活発な物質を含んでいる。視床下部の神経分泌細胞は電気的インパルスを導く能力を保っている。視床下部にある細胞体を刺激することによって活動電位が生じ、これが軸索の中を伝導してホルモン遊離のきっかけとなる。
Periventricular fibres(室周線維)Fibrae periventriculares
[A17_1_03_040]→室周線維は視床下部に起こって中脳被蓋および共、延髄の網様体を通り視床と視床下部をむすび、後方へ伸び背側縦束へつづく。
Inferior thalamic radiation(脚ワナ;脚係蹄)Radiatio inferior thalami
[A17_1_03_041]→下視床脚は視床放線のうちで最も発育が悪く、内側核と前頭葉底部、ならびに後核の一部と側頭葉外側面上面を結合する線維から成り、さらに内側膝状体核からでて横側頭回(聴覚中枢)に至る聴放線を含む。なお下視床放線には扁桃体、淡蒼球などから視床に至る線維も包含される。
Ansa lenticularis in corpus striatum(レンズ核ワナ;レンズ核係蹄)Ansa lenticularis in corpore striato
[A17_1_03_042]→レンズ核ワナの線維はおもに淡蒼球外節におこり、淡蒼球の腹側縁に沿って内方に走り、その内側縁ならびに内包の内側縁に沿って背方に転じてForel野に入り、ここで再び方向を背外方に転じて視床束(H1)として、視床外側腹側核(VL)に終止する線維束をいう。一方、淡蒼球内節におこる線維束は内包を横断してLuys体の背側に出てレンズ核束(H2)を形成し、内包に走ってForel野に入り、ここで反転してレンズ核ワナの線維束とともに視床束(H1)を構成する。視床束の線維は視床外側腹側核(VL)のほか、視床前腹側核(VL)のほか、視床前腹側核(VA)や視床正中中心核(CM)にも終止する。また、淡蒼球内節からForel野に達する線維の中には、さらに下行して橋の吻側被蓋部、とくに上小脳脚の周辺部のいわゆる脚橋被蓋核に終止するものがある。
Inferior thalamic radiation; Inferior thalamic peduncule(下視床脚)Radiatio inferior thalami; Pedunculus thalami inferior
[A17_1_03_043]→下視床脚は視床放線のうちで最も発育が悪く、内側核と前頭葉底部、ならびに後核の一部と側頭葉外側面上面を結合する線維から成り、さらに内側膝状体核からでて横側頭回(聴覚中枢)に至る聴放線を含む。なお下視床放線には扁桃体、淡蒼球などから視床に至る線維も包含される。
Fornix cerebrae(脳弓)Fornix
[A17_1_03_044]→脳弓は白い線維が帯状になった海馬体の主要な遠心性線維系を構成する。この中には投射線維と交連線維の両者が含まれる。これは海馬台皮質(海馬台前部、海馬台、前海馬台)と海馬の大錐体細胞の軸索からなり、海馬白質として側脳室表面に広がり、それらがまとまって海馬采を形成する。両側の海馬采は後方へ進むにつれて太くなり、海馬の後端に至って脳梁膨大の下を脳弓脚となって子を描いて上がると同時に両側の物が互いに近付いてくる。このあたりで多数の線維が反対側の脳弓に入る。すなわち交叉線維が薄く板状に広がって脳弓交連(海馬交連、または脳琴psalterium)を形成するがヒトでは発達が悪い。両側の脚は合して脳弓体となり脳梁の直下を前方に視床の吻側端まで行き、ここで再び線維束が左右に分かれ脳弓の前柱として室間孔から前交連の後ろまで腹方に曲がる。神経線維が薄い帯状になった海馬采は脳弓のほぼ全経過にわたって外側に位置しているが、吻側では脳弓の本体である脳弓前柱の中に混ざってしまう。脳弓線維の最大部分は前交連の尾側を交連後脳弓として下行し、残りは前交連の前を交連前脳弓となって走る。乳頭体では主として内側核に終止する。視床下部の吻側部で交連後脳弓から分かれた線維は、外側中隔核と、視床の前核群や外側背側核に終わる。その他、海馬台からの遠心線維は、前頭葉内側皮質、尾側帯状回、海馬傍回などに直接投射する。交連後脳弓線維の一部は乳頭体を越えてさらに尾方へ下り中脳被蓋に入る。
Crus of fornix(脳弓脚)Crus fornicis
[A17_1_03_045]→脳弓脚は左右がしだいに離れつつ下外側方に走り、ついで側脳室の下角に向かう。脳梁の下方で左右の脳弓脚の間には脳弓交連がある。
Body of fornix(脳弓体)Corpus fornicis
[A17_1_03_046]→脳弓体は脳梁の下にある脳梁の中央部で脳弓柱に続いて後走し、脳梁幹の下面と癒着しており、また左右が正中部で癒着している。脳梁幹の後部では再びこれと離れ、脳弓脚となる。
Taenia of fornix(脳弓ヒモ;脳弓紐)Taenia fornicis
[A17_1_03_047]→脳弓ヒモは脳弓の側方の薄い縁。側脳室の脈絡叢がつく。
Column of fornix(脳弓柱)Columna fornicis
[A17_1_03_048]→脳弓柱は没部と出部からでる。没部は左右の乳頭体から始まり、視床下部内を前上方に走り、前交連の後ろで出部に移行する。出部は大脳半球の正中断面で見える部分で、上行するとともにしだいに左右が互いに近づき、ついで後背側方に走り、前方は透明中隔板と癒着している。出部は脳梁幹の下で脳弓体に移行する。
Mammillothalamic fasciculus(乳頭体視床束;乳頭体視床路;乳頭視床束;乳頭視床路;ヴィック・ダジール束)Fasciculus mammillothalamicus
[A17_1_03_049]→乳頭視床束は乳頭体と視床前核を結ぶ神経線維束。主乳頭束として、乳頭被蓋束とともに乳頭体の背側よりでる。乳頭体内側核からおこる神経線維は同側の視床前核のうちでも前腹束核と前内側核に分布し、乳頭体外側核からおこる神経線維は両側の視床前背側核に分布するといわれる。また、乳頭視床束には視床前核から乳頭立ちに向かう神経線維も少数含まれるらしい。
Mammillotegmental fasciculus; Mammillotegmental tract(乳頭被蓋路)Fasciculus mammillotegmentalis; Tractus mamillotegmentalis
[A17_1_03_050]→乳頭体被蓋束は乳頭体内側核を中脳被蓋と結合して、視床下部が網様体に影響を及ぼすルートを形成している乳頭体被蓋束の線維は背側被蓋核に終止する。
Mamillary peduncle(乳頭体脚)Pedunculus corporis mamillaris にゅうとうたいきゃく
[A17_1_03_051]→乳頭体脚
(底部(乳頭体脚の))Pars basilaris(Pedunculus corporis mamillaris) [A17_1_03_052]→
(被蓋部(乳頭体脚の))Pars tegmentalis(Pedunculus corporis mamillaris) [A17_1_03_053]→
Olfactory tract(嗅索)Tractus olfactorius
[A17_1_03_054]→嗅索は前頭葉の下面で嗅球と嗅三角を結ぶ神経線維束で前有孔質に向かい、そこではっきり区別できる外側および内側嗅条に分かれる。嗅条の表面には薄い灰白質があってそれぞれ外側、内側嗅回とよばれる。外側嗅条と外側嗅回は前有孔質の外側縁を通って皮質の梨状葉皮質にいたり、その部分と、扁桃体の皮質内側核に終止する。
Habenulo-interpeduncular tract; Fasciculus retroflexus(反屈束)Tractus habenolointerpeduncularis; Fasciculus retroflexus
[A17_1_03_055]→手綱三角の内部には内側および外側手綱核がある。内側核は小細胞性で、外側核は大小の細胞を含む。これらの核は手綱の線維を受け、主として脚間核に向かう反屈束または手綱脚間路を出す。
(視床手綱核路)Tractus thalamohabenularis [A17_1_03_056]→
(嗅手綱路;嗅手綱核路)Tractus olfactohabenularis きゅうしゅもうろ[A17_1_03_057]→
Habenulo-interpeduncular tract; Fasciculus retroflexus(手綱核脚間路)Tractus habenolointerpeduncularis; Fasciculus retroflexus
[A17_1_03_058]→手綱三角の内部には内側および外側手綱核がある。内側核は小細胞性で、外側核は大小の細胞を含む。これらの核は手綱の線維を受け、主として脚間核に向かう反屈束または手綱脚間路を出す。
(嗅中脳路)Tractus olfactomesencephalicus きゅうちゅうのうろ [A17_1_03_059]→
(嗅海馬路)Tractus olfactohippocampicus きゅうかいばろ [A17_1_03_060]→
Stria terminalis(分界条)Stria terminalis
[A17_1_03_061]→分界条は背側視床と尾状核の間にある狭い白質で、主として扁桃体から起こり、脳弓と平行して走る。この線維の中には視索前野と視床下部と腹内側核でシナプス結合するものがある。
Lateral longitudinal stria(視床髄条;髄条)Stria longitudinalis lateralis
[A17_1_03_062]→外側縦条は帯状回の両外側縁の近くで、灰白質を伴って前後に走る細い縦条。嗅脳の一部。
Medial longitudinal stria(内側縦条)Stria longitudinalis medialis
[A17_1_03_063]→内側縦条は脳梁幹の表面で正中線の両端を、灰白質を伴って前後に走る細長い縦条。外側縦条とともに脳梁幹背面の灰白質を一部すなわち灰白質層、海馬の痕跡を形成する。嗅脳の一部。
Lateral longitudinal stria(外側縦条)Stria longitudinalis lateralis
[A17_1_03_064]→外側縦条は帯状回の両外側縁の近くで、灰白質を伴って前後に走る細い縦条。嗅脳の一部。
Subcallosal fasciculus(梁下束)Fasciculus subcallosus りょうかそく [A17_1_03_065]→
Brachium of superior colliculus(上丘腕)Brachium colliculi superioris
[A17_1_03_066]→上丘腕は上丘と外側膝状体とを連絡する視索線維束。
Brachium of inferior colliculus(下丘腕)Brachium colliculi inferioris
[A17_1_03_067]→下丘腕は下丘の外側にのびる隆起で、下丘と内側膝状体をつなぐ。上行性聴覚路の大部分を形成する。
(視床脊髄路)Tractus thalamospinalis ししょうせきずいろ [A17_1_03_068]→
Rubrospinal tract(赤核脊髄路)Tractus rubrospinalis
[A17_1_03_069]→『モナコフ束』ともよばれる。この伝導路の線維は、中脳被蓋の中心部にある卵円形の細胞集団である赤核からおこる。赤核は普通、吻側の小細胞群と尾側の大細胞群に分けられ、それらは動物によって大きさに差がある。赤核脊髄路は赤核の大細胞部から起こる。赤核脊髄路の線維は腹側被蓋交叉で完全に交叉し、脊髄各髄節を側索の皮質脊髄路の前側および一部それと混在して下行する。赤核脊髄路の線維は体部位局在性に構成されており、核の特定の部分の細胞はきめられた脊髄のレベルに選択的に投射する。頚髄へ投射する線維は赤核の背側および背内側部からおこり、一方腰仙髄に投射する線維は赤核の腹側および腹外側から起こる。胸髄は赤核の中間部から起こる線維を受ける。赤核脊髄路は①脊髄全長を下行する、②第Ⅴ層の外側半分、第Ⅵ層、および第Ⅶ層の背側部および中央部に終止する。赤核は大脳皮質と小脳から線維を受ける。“運動野”皮質からの皮質赤核路線維は赤核の小細胞部には両側性に、大細胞部には同側性に投射する。これらの投射線維はその起始終止とも体部位局在性に配列する。このシナプス結合を通して、皮質赤核路と赤核脊髄路とは共同して体部位局在性に配列した非錐体外路系伝導路として大脳皮質運動領と特定の脊髄レベルの間に存在する。赤核のあらゆる部分が上小脳脚を経てくる交叉性の小脳遠心線維を受ける。歯状核からの線維は赤核の前1/3に投射し、中位核(ヒトの球状核と栓状核に相当する)からの線維は体部位局在的に小脳皮質の部分と赤核の大細胞部とを関係づけている。赤核の細胞を刺激すると対側の屈筋のα運動ニューロンに興奮性シナプス後電位が発生し、また、伸筋のα運動ニューロンに抑制的シナプス後電位が発生する。赤核脊髄路の最も重要な機能は屈筋群の筋緊張の制御に関与することである。 Monakow, Constantin von (1853-1930) スイスの神経学者。大脳皮質の機能局在を明示し(Die Lolcalisation in Grosshirn u. der Abbau der Funktion durch kortical Herde, 1914)、モナコフ束(赤核脊髄路)を記述(Der rote Kern, die Haube u. die Regio hypothalamica bei einigen Saeugetieren und beim Menschen, Arb. Hirnanat. Inst. Zuerich, 1909, 3, 51-267; 1910, 4, 103-225)。
Rubroreticular tract(赤核網様体路)Tractus rubroreticularis せきかくもうようたいろ [A17_1_03_070]→
(赤核毛帯路;赤核絨帯路)Tractus rubrolemniscalis
[A17_1_03_071]→
Reticulospinal fibres; *Reticulospinal tract(網様体脊髄路;視蓋延髄路)Fibrae reticulospinales; *Tractus reticulospinales
[A17_1_03_072]→網様体脊髄路は、橋と延髄の網様体から脊髄に向かって下る様々な線維束をさす集合語。脳幹と脊髄において、この神経線維は内側縦束と関連して下行する。橋網様体脊髄路の線維は延髄に起始するものより多く、脊髄全長にわたって下行し、第Ⅷ層と第Ⅶ層隣接部に終止する。これらの網様体脊髄路線維は大部分が脊髄の1髄節以上にわたって側副枝を出し、これによって脊髄の異なったレベルでの協調活動に加わる可能性を示唆している。この網様体脊髄路の電気的刺激によって、体幹および四肢の骨格筋に分布する運動ニューロンに単シナプス性および多シナプス性の活動が起こる。直接的な影響は体幹の、特に頚部の筋に最も強い。
Predorsal bundle(背前束;視蓋延髄および視蓋脊髄路)Fasciculus predorsalis; Tractus tectobulbaris et tectospinalis はいぜんそく;しがいえんずいおよびしがいせきずいろ
[A17_1_03_072_1]→背前束は上丘深層灰白質におこり、中心灰白質縁と同心円状に中脳網様体を横切って背側被蓋交叉を経て対側にわたり、内側縦束の腹方位置して、これからは方向を尾方に転じてほぼ直線的に下行して頚髄後柱基部や中間帯に終止する。前者を視蓋被蓋路、後者を視蓋脊髄路とよぶ。上丘を破壊してナウタ法でみると、背前束から内側腹オリーブ核へ終止性変性線維が認められることから、視蓋オリーブ路が示唆されている。なお、上丘におこり背前束を通って外眼筋支配核に終止する神経線維があるとされている。
Tectospinal tract(視蓋脊髄路)Tractus tectospinalis
[A17_1_03_073]→視蓋脊髄路は主として中脳の上丘(第4および第6層)および一部は下丘から出て、すぐに背側被蓋交叉で交叉し、反対側の脳幹の正中部の知覚で内側縦束の腹側を下り(背前束)、脊髄では前索の前正中裂に近い部分を下行し、頚髄、ことに上位頚髄の前角の腹内側部から灰白質に入り、中間質外側部およびその付近に分布するが、直接に前角細胞に終わるものはない。これは視覚、一部は聴覚刺激に応じた頭部、上肢の反射的姿勢運動による。
Medial tectospinal tract(内側視蓋脊髄路)Tractus tectospinalis medialis [A17_1_03_074]→
Lateral tectospinal tract(外側視蓋脊髄路)Tractus tectospinalis lateralis [A17_1_03_075]→
(被蓋路交叉)Decussatio tractuum tegmenti [A17_1_03_076]→
Spinotectal tract(脊髄視蓋路)Tractus spinotectalis
[A17_1_03_077]→脊髄視蓋路は上丘深層へ投射するがその数は多くない。脊髄視蓋路の起始細胞は後角の第Ⅰ層と第Ⅴ層に存在している。この交叉性伝導路の神経戦は脊髄視床路系に密接して脊髄の前外側部を上行するが、中脳の高さで上丘の中間層と深層と中心灰白質の外側部に投射する。上丘への体性感覚入力は脊髄の第Ⅳ層の細胞からも幾分かは起こるが、主な入力は楔状束核および三叉神経脊髄路核のすべての部分から起こる。これらの入力は体位部位的局在的に構成されており、頭部からの近くは上丘の上部に終わる。上丘は、①主に視覚入力を受ける浅層と②いろいろな多感覚性の入力を受ける深層の2つに区分けされる。浅層は深層とは明らかに異なる投射様式を示すが、この浅層もまた深層に投射する。
Vestibulospinal tract(前庭脊髄路;前庭神経脊髄路;ヘルド束)Tractus vestibulospinalis; Tractus spinalis nervi vestibularis ぜんていせきずいろ;へるどそく
[A17_1_03_078]→
Posterior longitudinal fasciculus; Dorsal longitudinal fasciculus(後縦束;背側縦束)Fasciculus longitudinalis posterior; Fasciculus longitudinalis dorsalis
[A17_1_03_079]→背側縦束は中脳から延髄にかけて中心灰白質の腹内側部にみれれる小さい神経線維束で、細い有髄線維を含む。上行性および下行性の比較的短い神経線維の連鎖であり、吻側では視床下部の室周線維に連絡する。自律性または内臓性情報の中枢伝導系の一つとされる。
Medial longitudinal fasciculus; MLF(内側縦束)Fasciculus longitudinalis medialis
[A17_1_03_080]→前索の後部には脳幹のいろいろなレベルにある種々な神経核からでる複雑な下行線維束がある。この複雑な神経線維束は内側縦束として知られている。この神経束の脊髄部は同じ名称で呼ばれる脳幹にある伝導路の一部にすぎない。内側縦束の上行線維は主として前庭神経内側核および上核から起こり、同側性および対側性に主として外眼筋支配の神経核(外転、滑車、動眼神経核)に投射する。内側核からの上行線維は主に交叉をし、両側の外転神経核と左右の動眼神経核に非対称性に終わるが、滑車神経核へは対側性に投射する。上核の中心部にある大形細胞は非交叉性上行線維を内側縦束に出し、これは滑車神経核および動眼神経核に終わる。同核の周辺部にある周辺部にある小型細胞は交叉性の腹側被蓋束(内側縦束の外側にある)を経て動眼神経核に投射するが、これは主として対側の上直筋を支配する細胞に作用する。生理学的には、前庭神経核から外眼筋支配核から外眼筋支配核への上行性投射のうち、交叉性線維は促進的に働くが、非交叉性線維は抑制的に働く。内側縦束にはこのほかに、左右の外転神経核の間にある神経細胞から起こり、交叉して上行し、動眼神経核の内側直筋支配部に終わる明瞭な線維が含まれる。この経路は一方の外転神経核の活動を対側の動眼神経核内側直筋支配部へ連絡する物で、外側視の場合に、外側直筋が収縮すると同時に対側の内側直筋が共同して収縮するための神経機構を形成している。内側縦束の上行線維の一部は、動眼神経核を回ってCajal間質核に終わる。これは内側縦束内にうまっている小さい神経細胞群である。前庭神経内側核は対側性に間質核へ投射するが、上核は同側性に終わる。前庭神経二次線維は両側性に視床の中継核へ投射し、その数は中等度で、後外側腹側核に終止する。前庭からの入力を受ける視床の細胞は体性感覚情報にも対応するが、これは視床には特定の前庭感覚中継核がないことを示唆している。
Mesencephalic tract of trigeminal nerve(三叉神経中脳路)Tractus mesencephalicus nervi trigemini
[A17_1_03_081]→三叉神経中脳路は中脳中心灰白質に沿って位置し、単一の知覚神経からなり、その起点の細胞は三叉神経中脳路核の細胞体からの主な突起は鎌状をした三叉神経中脳路を作り、これは三叉神経運動核の高さまで下行し、側副枝を運動核に送るが、大部分は運動根の一部として脳外に出る。
Spinal tract of trigeminal nerve; Spinal tract of CN V(三叉神経脊髄路)Tractus spinalis nervi trigemini
[A17_1_03_082]→三叉神経脊髄路は延髄から橋の横断面上にコンマ状に明確に認められる線維束。脊髄路核の各部の第Ⅰ層と第Ⅲ層細胞から起こる。これらのうち、下部と中間部からの線維は同側性であるが、上部からのものは両側性に下行し、後柱に入る感覚性情報を調節し、種々の反射に関係し、さらに三叉神経支配の受容器と脊髄の体性および内臓性効果器を連絡している。
Medullary striae of fourth ventricle(第四脳室髄条;髄条)Striae medullares ventriculi quarti
[A17_1_03_083]→第四脳室髄条は後脳部と髄脳部の境界部の脳室底には数本の線維が正中溝を横切って横走する線維の束。下小脳脚まで追跡できる。髄条は、次の諸部分から起こる線維より成る複雑な線維束である。すなわち、①中隔核、②外側視索前野、③視床前核群である。海馬体および扁桃体複合核からの線維は中隔核に投射する。
Trapezoid body(台形体;梯形体)Corpus trapezoideum
[A17_1_03_084]→台形体は橋下位の高さで蝸牛神経腹側核および一部台形体背側核(上オリーブ核)から出て橋被蓋の背側部を横走し対側に向かう線維の総称(もしくはこれらの線維で構成された部位)。これらの線維は交叉の後、前背側方に進み、外側毛帯に加わる。
Solitary tract(孤束)Tractus solitarius
[A17_1_03_085]→孤束は被蓋背側部を縦走する細いが明瞭な線維束。孤束核で囲まれ、閂の下方で左右のものが中心管の背側で交叉し、ある距離上を下り頚髄上部へと入る。迷走神経、舌咽神経および顔面神経(中間神経)に由来する内臓求心性線維によって形成されている。下の前2/3の味覚を伝道する神経(鼓索神経)と下の後1/3部の味覚伝導神経(舌咽神経)は主として孤束核の吻側に部に終わる。迷走神経が入る高さとその尾方の孤束は主として一般内臓求心性線維を含み、大部分は迷走神経からくる。
Spinobulbar fibres; Spinobulbar tract(脊髄延髄路)Fibrae spinobulbares; Tractus spinobulbares [A17_1_03_086]→
(内側毛帯路;内側絨帯路)Tractus lemnisci medialis [A17_1_03_087]→
Spinothalamic tract(脊髄視床路)Tractus spinothalamicus せきずいししょうろ
[A17_1_03_088]→この伝導路は前脊髄視床路と密接に関係しているが、痛覚や温度覚に関係するインパルスを伝達するため、臨床医学上はるかに重要である。この経路の線維はより集中化され、視床に直接投射する長い線維を含んでいる。前脊髄視床路の起始細胞に関する知見は外側脊髄視床路にもあてはまる。主として第Ⅰ層、Ⅳ層および第Ⅴ層の細胞が、白前交連で交叉して反対側の側索を外側脊髄視床路として上行する軸索の大部分を出している。この伝導路の線維は斜めに反対側へ横断するが、普通一髄節内で行われる。この伝導路はまた前脊髄視床路と類似した配列で体部位局在的に構成されており、また前脊髄小脳路の内側に位置している。痛覚と温度覚に関係する神経線維は不完全に分離している。温度覚に関係する線維は痛覚に関係する線維の後側に位置する傾向がある。脳幹にあってはこの伝導路は網様体に枝を出すが、主幹は視床の後外側腹側核尾部(VPLc)に終わる。
Anterior spinothalamic tract; Ventral spinothalamic tract(前脊髄視床路)Tractus spinothalamicus anterior えんずいししょうろ
[A17_1_03_089]→脊髄視床路は後索の線維とちがって、脊髄内のニューロンから始まる。脊髄灰白質内から始まる線維は交叉して視床まで上行する。この伝導路は脊髄内で交叉し視床に達するという、古くから知られた経路であるが、その起始細胞はごく最近になって確定された。脊髄視床路の起始細胞は次のようにして証明された。①生理学的には、視床の特殊感覚中継核の逆行性刺激、②解剖学的には、西洋ワサビ過酸化酵素horseradish peroxidase (HRP)を用いた逆行性軸索流の追跡によった。これらのデータは、脊髄視床路の線維が脊髄では対側性に主に第Ⅰ、Ⅳおよび第Ⅴ層の細胞からでることを裏付けているもっとも、いくらかの線維は第Ⅵ層と第Ⅶ層の細胞から起こっている。霊長類の動物で、大量のHRPを注入すると、多数の脊髄視床路のニューロンが反対側の下位腰髄に見出される。脊髄の各領域では同側性に標識された脊髄視床路のニューロンは全体の約10%を占めている。脊髄視床路を形成するニューロンは異なった髄節や灰白層では大きさ、形、数量が変わる不均質な細胞分布でできている。後角内の一時間核線維の神経終末とこれと関わるシナプス結合に関する詳細は不明であるが、求心性線維が第Ⅳ層と第Ⅴ層の細胞の樹状突起と接触していることは考えられることである。脊髄視床路は白前交連で交叉するが、その交叉はいくつかの髄節わたり、また対側性に上行する。前脊髄小脳路を形成する線維は前索と前外側索を上行し、また、体部位局在的に配置されているので、仙髄と腰髄からはじまる線維は最も外側で、胸髄と頚髄からのものは最も内側を占める。前脊髄視床路は、一部の線維かまたは側枝が網様体の核に投射しているから、延髄の高さで大きさが小さくなる。この伝導路を構成する脊髄視床路線維は橋および中脳で内側毛帯と密接に関連をもっている。中のレベルでは前脊髄視床路は2つの構成成分から成る。外側部分の線維は大きくて視床後核と後外側腹側核尾部(VPLc)視床核に終止し、内側の線維は中心灰白質と両側性に髄板内核とに投射する。前脊髄視床路の線維は”軽い触覚”の感覚に関係するインパルスを伝達する。この感覚は毛のないところの皮膚を羽毛や綿辺でさすることで起こされる。前脊髄視床路の損傷は、たとえあるとしても、非常に軽い障害しか起こさない。これは触覚が、後索によっても伝達されるからである
Lateral spinothalamic tract(外側脊髄視床路)Tractus spinothalamicus lateralis
[A17_1_03_090]→この伝導路は前脊髄視床路と密接に関係しているが、痛覚や温度覚に関係するインパルスを伝達するため、臨床医学上はるかに重要である。この経路の線維はより集中化され、視床に直接投射する長い線維を含んでいる。前脊髄視床路の起始細胞に関する知見は外側脊髄視床路にもあてはまる。主として第Ⅰ層、Ⅳ層および第Ⅴ層の細胞が、白前交連で交叉して反対側の側索を外側脊髄視床路として上行する軸索の大部分を出している。この伝導路の線維は斜めに反対側へ横断するが、普通一髄節内で行われる。この伝導路はまた前脊髄視床路と類似した配列で体部位局在的に構成されており、また前脊髄小脳路の内側に位置している。痛覚と温度覚に関係する神経線維は不完全に分離している。温度覚に関係する線維は痛覚に関係する線維の後側に位置する傾向がある。脳幹にあってはこの伝導路は網様体に枝を出すが、主幹は視床の後外側腹側核尾部(VPLc)に終わる。
Bulbothalamic Tract(延髄視床路)Tractus bulbothalamicus [A17_1_03_091]→
(聴外側絨帯路;外側絨帯路)Tractus lemnisci lateralis (acustici) [A17_1_03_092]→
(髄条繊維)Fibrae striatum medullarium [A17_1_03_093]→
Decussation of medial lemniscus; Sensory decussation(毛帯交叉;絨帯交叉)Decussatio lemnisci medialis; Decussatio lemniscorum
[A17_1_03_094]→内側毛帯は感覚性の神経路で、交叉線維でできる。この交叉を内側毛帯交叉(感覚交叉)という。
Medial lemniscus(内側毛帯;内側絨帯)Lemniscus medialis
[A17_1_03_095]→延髄の後索(薄束および楔状束)を通過する伝導路は圧覚と触覚や固有知覚の興奮を後索核(薄束核および楔状束核)や視床を経て大脳皮質に伝達する神経路である。薄束核および楔状束核から起こる二次ニューロンは延髄視床路(内側毛帯)となり正中縫線近く延髄の中心を通り上行する。橋にはいると外側に広がり、橋核の背側縁を越えて上行する扁平な帯になる。中脳内では、黒質の背側縁を越えて赤核で外側に移る。内側膝状体まで内側を通り視床の後腹側核に入り、そこで終わる。視床を出た第3ニューロンの線維は、上行して大脳皮質におもむく。内側毛帯系は脊髄から上行する識別性感覚路の最初の一環を形成するのは後根を通って入ってくる太い有髄神経の枝であり、後索を上行する。後索を上行するこれらの神経線維は身体部位対応配列を示す。すなわち、仙骨神経根や腰神経塊を通って入ってくる上行枝は後索の内側部を占めて薄束を形成する。一方、頚神経根を通って入ってくる上行枝は後索の外側部を占めて楔状束を形成する。また、胸神経根を通って入ってくる少数の上行枝は、薄束と楔状束との間に位置する。薄束と楔状束は、延髄の尾側端でそれぞれ対応する神経核、すなわち、薄束核と[内側]楔状束核に終止する。ある後根が支配する皮膚領域は、同時にその後根の上下の後根からも支配されている。このように一定の皮膚領域を支配する隣接後根の神経線維群は、後根から後索、さらに後索核へと向かう経過のうちに、一つの神経束にまとまる。このような集束の結果として、隣接する皮節(dermatome)間の重なり合いは解消されるのであるが(一つの皮節からの情報を伝道する神経線維が集合して一つにまとまる)、後索で最初にみられたような層構造は次第に不明瞭になる。薄束核の背側部と[内側]楔状束核の背側部にはニューロンが幾つかの小群をつくって分布する。超す悪を上行する神経線維の中で、四肢の遠位部を支配するものがこれらのニューロン小群に終止して身体部位対応配列を示す。後索核の腹側部と吻側部では身体部位対応配列はあまり精細でない。後索には後核固有核から起こる内在性神経線維も含まれている。これらの内在性神経線維は後索核の腹側部と吻側部に終止する。その他、後側索(側索後部)を上行する神経線維は両側の後索核の腹側部と吻側部、およびZ群(group Z)に終止する。Z群は薄束核の吻側端に位置するニューロン群であり、下肢の筋からの入力を視床に中継する。上枝からの固有感覚性入力を中継するのは外側楔状束核である。この核から起こる投射神経線維は主として下小脳脚を通って小脳へ入る。後索核の腹側部と吻側部から起こる投射神経線維は、後索核背側部(ニューロンの小群の集合から成る)から起こる投射神経線維に比べて、分布範囲が広い。すなわち、前者も後者も反対側の視床へ向かうのであるが、前者はさらに小脳や下オリーブ核に投射し、脊髄の後角へ向かうものもある。薄束核と[内側]楔状束核からは内弓状線維が起こり、正中部で交叉してのち、内側毛帯を形成して上行し、視床の後外側腹側核、後核群、内側膝状体大細胞部、および不確帯に終止する。後索核から後外側腹側核への投射は”核と殻(core-and-shell)”の様式を示す。すなわち、後索核背側部から起こる皮膚感覚の投射線維は後外側腹側核の中心部(すなわちcoreの部分)に終止し、後索核腹側部と吻側部から起こる固有感覚の投射線維は後外側腹側核の辺縁部(すわなち、shellの部分)に終止する。内側毛帯線維は後外側腹側核において一連の平行な層板をなして終止する。これらの層板は核のcoreの部分とshellの部分を通じて前後方向に伸びており、それぞれの層板が身体の特定の部位に対応している。また、各層板の前後軸に沿って、種々の感覚要素に対応する投射線維の終末が次々と配列分布する。
Lateral lemniscus(脊髄毛帯)Lemniscus lateralis
[A17_1_03_096]→外側毛帯は中脳まで上行し、大部分の線維が下丘に終わる。背側および腹側蝸牛神経核からの線維は、背側、中間および腹側聴条として対側に向かい、多くは対側の台形体背側核におわるが一部はそのまま上行する。この上行する線維と同側の台形体背側核から出て上行する線維が一緒になって外側毛帯を形成する。外側毛帯は橋の高さで内側毛帯(系)の背外側の位置を占めて上行し、大部分は下丘に終わるが、一部は途中下丘のすぐ腹側に存在する外側毛帯核におわる。
Trigeminal lemniscus; Trigeminothalamic tract(三叉神経毛帯)Lemniscus trigeminalis; Tractus trigeminothalamicus
[A17_1_03_097]→解剖学用語としては、三叉神経脊髄路核、三叉神経主知覚核におこり視床におわる伝導路をいうが、三叉神経主知覚核の腹側2/3におこり交叉して視床に上行する線維束のみを指す場合がある。三叉神経視床路は主に脊髄路核の下部と中間部の第Ⅰおよび第Ⅳ層細胞から起こる。これらの細胞の軸索は網様体中を腹内側方に向かい、正中線で交叉して対側の内側毛帯のすぐ近くをこれに沿って走り、視床の後内側腹側核の細胞に選択的に終わる。
Lateral lemniscus(外側毛帯;外側絨帯)Lemniscus lateralis
[A17_1_03_098]→外側毛帯は中脳まで上行し、大部分の線維が下丘に終わる。背側および腹側蝸牛神経核からの線維は、背側、中間および腹側聴条として対側に向かい、多くは対側の台形体背側核におわるが一部はそのまま上行する。この上行する線維と同側の台形体背側核から出て上行する線維が一緒になって外側毛帯を形成する。外側毛帯は橋の高さで内側毛帯(系)の背外側の位置を占めて上行し、大部分は下丘に終わるが、一部は途中下丘のすぐ腹側に存在する外側毛帯核におわる。
Inferior cerebellar peduncle(下小脳脚;髄小脳脚;索状体)Pedunculus cerebellaris inferior
[A17_1_03_099]→下小脳脚は複合線維束であり、脊髄と延髄の細胞群から起こり小脳へ投射する線維で構成されている。この線維は副楔状束核の外側、および三叉神経脊髄路の背側の延髄後外側縁に沿って集まり下小脳脚に入る。索状体という外側の大きな束と索状体傍体という内側の小さな束とからなる脊髄ニューロンと延髄中継核からの線維の複合体である。この線維束は上部延髄ではさらに多くの線維が加わることによって急速に増大し、小脳に入る。交叉性のオリーブ小脳路の線維は下小脳脚の最大の構成成分をなしている。下小脳脚を通る求心路には後脊髄小脳路、副楔状束核小脳路、オリーブ小脳路などがある。前庭小脳路、網様体小脳路、三叉神経小脳路などの求心路や小脳皮質前庭路、室頂核前庭路(鈎状束)などの遠心路は下小脳脚内側部、すなわちMeynertのIAK(innere Abteilung des uteren Kleinhirnstiels, Corpus juxtarestifome)とよばれるところを通る。下小脳脚を経由して小脳にインパルスを中継するその他の延髄の核は、①延髄の外側網様核、②副楔状束核、③傍正中網様核、④弓状核、⑤舌下神経周囲核である。外側網様核と副楔状束核からの投射は非交叉性であるが、他の延髄中継核からくるものは交叉性、非交叉性の両方がある。非交叉性の後脊髄小脳路もこの脚を経由して小脳に投射する。
Middle cerebellar peduncle(中小脳脚;橋小脳脚;橋腕)Pedunculus cerebellaris medius; Brachium pontis
[A17_1_03_100]→中小脳脚(橋腕)は3対ある小脳脚のうち最大のもので、主として橋核から起始する線維からなり、橋底の正中線を越えて対側の背側に移り太い束となって橋被蓋の外側を乗り越えて小脳にはいる。少数の対側へ移らない線維もある。少数の側副線維が小脳核に達している以外ほとんどが橋小脳路線維からできている。
Superior cerebellar peduncle(上小脳脚;小脳大脳脚;結合腕)Pedunculus cerebellaris superior; Brachium conjunctivum; Crus cerebellocerebrale
[A17_1_03_101]→上小脳脚(結合腕Brachium conjunctivum)は主として小脳を出る線維からなる。その主体をなす線維は小脳視床路と小脳赤核路である。これらは主として歯状核から出て、腹内側方に進んで深部に入り、中脳下半で大部分交叉し、上小脳脚交叉(結合腕交叉)を作り、反対側の中脳被蓋を上行し、一部は赤核に終わるが(小脳赤核路)、一部はさらに視床の前外側腹側核に至る(小脳視床路)。なお上小脳脚の表面を前脊髄小脳路が逆行して小脳に入り、主としてその前葉に分布する。また鈎状束は室頂核から出て大部分交叉し、上小脳脚の背外側をへて鈎状に曲がり、下小脳脚内側部の上部に来て前庭神経各核にならびに橋、延髄の網様体内側部に分布する。
Cerebello-rumbro-thalamic fibers(小脳赤核視床路;小脳赤核視床線維)Tractus cerebello-rubro-thalamicus しょうのうせきかくししょうろ;しょうのうせきかくししょうせんい [A17_1_03_101_1]→
Cerebellorubral tracts; Cerebellorubral fibers(小脳赤核路;小脳赤核線維)Tractus cerebellorubralis しょうのうせきかくせんい;しょうのうせきかくせんい
[A17_1_03_102]→ 中脳の上部で、被蓋のほぼ中央にある神経核である。核は断面で直径約5mmの卵円形を呈し、新鮮な脳ではやや赤みをおびる。このように淡赤色を呈するのは、核が鉄を含み毛細血管に富むためといわれる。 赤核は大脳・小脳などからの線維(入力)を受ける中継核で、とくに大部分の線維を上小脳脚を経て小脳から受けている(小脳赤核線維)。赤核から起こる線維(出力)は主として交叉して(被蓋交叉)下行し、脳神経の運動核・網様体・脊髄前角に至る(赤核脊髄路)。ヒトでは、網様体を経て脊髄に達するといわれる(赤核網様体路)。(解剖学講義)
Decussation of superior cerebellar peduncles(上小脳脚交叉;小脳大脳脚交叉;結合腕交叉)Decussatio pedunculorum cerebellarium superiorum; Decussatio crurum cerebellocerebralium
[A17_1_03_103]→上小脳脚は小脳核から出て初めは表面を走るが、下丘の高さで奥に入り交叉して一部は対側の赤核に終わり、一部は視床に至る。この交叉を上小脳脚交叉という。
Cerebellothalamic fibers(小脳視床路;小脳視床線維)Fibrae cerebellothalamicae しょうのうししょうろ;しょうのうししょうせんい
[A17_1_03_104]→上小脳脚の線維で視床の外側腹側核にはいる。
Rubrothalamic tract(赤核視床路)Tractus rubro-thalamicus せきかくししょうろ
[A17_1_03_105]→
Fastigiobular tract(室頂核延髄路;小脳延髄路)Tractus fastigiobulbaris しつちょうかくえんずいろ、しつちょうえんずいろ;しょうのうえんずいろ
[A17_1_03_106]→室頂核延髄路は左右の室頂核(視蓋核)から発する線維束で、下小脳脚(索状体)内側を通って小脳を通過し、その線維は前庭神経核および延髄内の他の神経核に分枝する。交叉神経の多くは、腹側に曲がる前に上小脳脚背側面上で環状になり、Russell鈎状束を形成する。
Cerebellovestibular tract; Corticovestibular tract(小脳皮質前庭神経核路;小脳前庭神経核路;小脳前庭核路;皮質前庭神経核路)Tractus cerebellovestibularis; Tractus corticovestibularis しょうのうひしつぜんていしんけいかくろ;しょうのうぜんtねいしんけいかくろ;しょうのうぜんていかくろ;ひしつぜんていしんけい
[A17_1_03_107]→小脳皮質の特定の部位から前庭神経核へ直接の投射がある。これらの線維は小脳虫部と前庭小脳(片葉小節小葉)にあるPurkinje細胞の軸索である。このように、前庭神経核は室頂核から両側性に、また特定の小胞皮質部から同側性に線維を受ける。小脳虫部からの直接投射は同側の前庭神経外側核と下核の背側部に体部位的局在性をもって終わり、GABAを伝達物質としている。前葉虫部を刺激すると、前庭神経外側核の細胞で単シナプス抑制を起こす。前庭一次および二次線胃を受ける小脳の部分を「前庭小脳」といい、片葉小節小葉の他に虫部垂の腹側部も含まれる。「前庭小脳」のすべての部分から前庭神経核への投射がある。すなわち、片葉からは前庭神経上核と内側核へ、小節と虫部垂からは上核、内側核および下核へ投射があり、これらはすべて同側性である。
Olivocerebellar tract(オリーブ核小脳路;オリーブ小脳路)Tractus olivocerebellaris おりーぶかくしょうのうろ;おりーぶしょうのうろ
[A17_1_03_108]→オリーブ核小脳路は下オリーブ核から小脳に投射する経路である。交叉性で下小脳脚を通り小脳皮質のほぼ前域とすべての小脳核に投射する。皮質では登上線維となりPurkinje細胞と結合する。下オリーブ核の主オリーブ核と背側および内側副オリーブ核内の尾状な領域と小脳皮質との間に明瞭な投射の局在関係が認められる。大きく分けて虫部皮質は内側副オリーブ核から、虫部傍皮質は内側および背側副オリーブ核から、半球皮質は主オリーブ核からの線維を受ける。
(歯状核オリーブ路)Tractus dentatoolivaris しじょうかくおりーぶろ [A17_1_03_109]→
Posterior spinocerebellar tract; Dorsal spinocerebellar tract(オリーブ小脳路)Tractus spinocerebellaris posterior
[A17_1_03_110]→後脊髄小脳路は胸髄核から出て交叉せずにすぐ側索周辺部の背側部を上行し、下小脳脚を通って同側の小脳の前葉、一部は虫部錐体、虫部垂などの皮膚に達する。
Anterior spinocerebellar tract; Ventral spinocerebellar tract(前脊髄小脳路)Tractus spinocerebellaris anterior
[A17_1_03_111]→前脊髄小脳路は発育が悪く、腰髄の後角底および中間質外側部などの神経細胞から出て、主として白交連で交叉し、反対側の側索周辺部の腹側部を上行し、延髄の側索をへて橋に達し、背側方に進んで、上小脳脚の表面を逆行し、ついで再び交叉し、主として小脳前葉の皮膚に入る。前および後脊髄小脳路は下半身からの深部知覚を伝える。これに対して、上肢、頚部、体感上部などからの深部知覚は副楔状束核小脳路によって伝えられる。
Posterior spinocerebellar tract; Dorsal spinocerebellar tract(後脊髄小脳路)Tractus spinocerebellaris posterior
[A17_1_03_112]→後脊髄小脳路は胸髄核から出て交叉せずにすぐ側索周辺部の背側部を上行し、下小脳脚を通って同側の小脳の前葉、一部は虫部錐体、虫部垂などの皮膚に達する。脊髄の側索後外側辺縁部を上行するこの非交叉性の伝導路は胸髄核の大細胞から起始する。後根の求心性線維は直接に、または後索を上下行した後に胸髄核に終わる。胸髄核の大細胞は太い線維を出し、これは側索の後外側部(すなわち皮質脊髄路の外側)に入り、上行する。延髄にあってはこの伝導路の線維は下小脳脚に組み込まれ、小脳に入って同側性に虫部の吻側と尾側に終わる。虫部全部では線維は第Ⅰ小葉から第Ⅳ小葉に終わり、後部では主として虫部錐体と正中傍小葉に終わる。胸髄核は第三胸髄から尾方には存在しないから、尾方の髄節からの後根線維はまず後索内を上位の胸髄まで伝達され、それから胸髄核の細胞へ伝えられる。後脊髄小脳路を経由して小脳へ中継されるインパルスは伸展受容器である筋紡錘やGolgi腱器官および触圧覚受容器から起こる。胸髄核のニューロンは主としてⅠa群、Ⅰb群およびⅡ群の求心線維を経由する単シナプス性興奮を受ける。Ⅰ群の求心性線維と胸髄核の間のシナプス結合では高頻度のインパルスの伝達が行われる。一部の外受容器由来のインパルスもまた後脊髄小脳路を経由して伝達される。これらは皮膚の触覚と圧覚の受容器およびゆっくり反応する圧受容器に関係する。後脊髄小脳路は脊髄レベルおよび小脳の終始部において体部位局在性に配列されている。伝導路によって伝達されるインパルスは意識のレベルに達することはない。これらの伝導路によって伝達されるインパルスは姿勢とここの四肢筋の運動の細かい協調作用に役立っている。
Gracile fasciculus(薄束;内側部(後索の))Fasciculus gracilis; Pars medialis funiculus posterior
[A17_1_03_113]→薄束はゴル束ともよばれる。楔状束(ブルダッハ束)とともに脊髄後索をなす。両側の後索は胸髄上部と頚髄において後中間中隔によって2分される。この中隔はおよそ第六胸髄の高さで認められ、薄束(内側)と楔状束(外側)を分ける。薄束は脊髄全長にわたって存在し、仙髄部、腰髄部、下位6胸髄部の後根由来の長い上行枝を含む。薄束は後索の内側部にある。体の下半分から線維を含む。触覚と深部知覚を伝える。Goll, Frindrich (1829-1903)スイスの解剖学者、チューリヒ大学の教授。脊髄後索の内側部(薄束)について1860年に記述(「Beitrage zur feineren Anatomie・・・」;, Denk. Medchir. Ges. Kanton Zurich, 1860, 130-171).
Cuneate fasciculus(楔状束;外側部(後索の))Fasciculus cuneatus; Pars lateralis funiculus posterior
[A17_1_03_114]→楔状束はブルダッハ束ともよばれる。楔状束は最初、およそ第六胸髄の高さで出現し、上位の6胸神経と前頚神経の後根の長い上行枝を含む。薄束と楔状束の神経線維は同側を上行し、後索の延髄中継核、すなわち薄束核と楔状束核に終わる。後索系には2部があり、薄束(Goll束)および楔状束(Burdach束)として脊髄の後索を上行する。これらの線維束は太い後根線維の直接の続きであって、延髄の後索核にまで達してシナプス結合する。後索系は主として四肢から起こる線維からなり、系統発生的に新しくて、ヒトでもっとも発達している。ヒトではこの線維の長さは長いもので約150cmである。楔状束は後索の外側部。身体の上半分から起こる線維を含む。ドイツの解剖・生理学者Karl Friedrich Burdach (1776-1847)の名を冠する。以前に同部についての報告はあったが、ブルダッハの正確な報告で知られるようになった。
Internal arcuate fibres(内弓状線維)Fibrae arcuatae internae
[A17_1_03_115]→内弓状線維は薄束核と楔状束核から起こる有髄神経線維は内弓状線維となり中心灰白質をまわって前内方へ移動する。これらの線維は完全に交叉し、内側毛帯とよばれる明瞭な上行束を形成する。
Anterior external arcuate fibres of medulla; Ventral external arcuate fibres of medulla(前外弓状線維)Fibrae arcuatae externae anteriores
[A17_1_03_116]→延髄の前外弓状線維は延髄基部の弓状核から出て延髄の外側表面に沿って走っているもの。後外側弓状線維とともに下小脳脚の索状体として小脳にはいる。
Posterior external arcuate fibres; Dorsal external arcuate fibres(後外弓状線維)Fibrae arcuatae externae posteriores
[A17_1_03_117]→後外弓状線維は弓状核の外側部よりでて下小脳脚にいたる線維。内耳神経(Ⅷ)核領域ではこの線維は後脊髄小脳路の代わりをなす。この部分では胸髄核を欠く。
Gelatinous substance; Dorsolateral fasciculus(膠様質;後外側束(後角尖の))Substantia gelatinosa; Fasciculus dorsolateralis [A17_1_03_118]→
Interfascicular fasciculus(束間束;半月束;コンマ束;半円束;下行部;コンマ状部(後索の))Fasciculus interfascicularis; Fasciculus semilunaris; Pars descendens funiculus posterior
[A17_1_03_119]→後索の主体を構成する薄束と楔状束の間に存在するコンマ形の領域をコンマ野comma fieldとよび、この部を形成する線維群をコンマ束comma tractという。この神経束の存在はWestphal (1880)によって最初に記載されたが、Schultze(1883)がコンマ束の名称を使ったことから後年Schultzesches Kommaの用語が広く使われるようになった。かれはC4とC5のレベル後根を横断したとき、手術巣から約2.5cm尾側でコンマ上の領域に変性繊維の集団を出現することを記載した。コンマ束はその位置から束間束または半月束ともよばれる。典型的なコンマ状を呈する変性線維の束は、頚髄に侵入する後根を切断したとき、胸髄上部の後索で観察することができる。しかしその時もコンマ束の形状と出現位置は正確には一致しない。コンマ状を尾方にに辿るとその形が変化し、コンマの頭の部分だけになり、尾の部分がなくなる。胸髄下半分以下で後根を切断しても、腰仙髄への後索でコンマ束を認めることはできない。これらのことを総称して、この固有束は頚神経と第1~7胸神経の後根由来の付帯的な下行枝によって構成される神経束とであると考えられる。胸髄下半分とそれ以下の後根から進入する線維の下行枝は後索内では分散して走行し、コンマ束となってまとまることはない。コンマ束は頚髄および胸部上半部に親友する後根線維の下行性分枝からなり、胸髄中央部以下に入る後根線維の下行枝はコンマ野に入らず、分散して下行枝、まとまりを示さないと考えられる。内在性の介在ニューロンの軸索の参加は完全には否定できないが、たとえあってもその数は少ない。T7以下で後根を切断してもコンマ束線維の変性は起こらない。後根線維の下行性分枝はコンマ区束、または中隔縁束、とくに卵円野領域を主として下行するとみられる。人ではC5からC6後根線維の下行枝はT12レベルまで追跡でき、C8の切断でもT12までたどれる。T4での切断では仙髄まで変性線維を追うことができる。これらの線維は後柱および後交連の領域に進入し、終末する。
Septomarginal fasciculus(中隔縁束;内側視蓋脊髄路)Fasciculus septomarginalis
[A17_1_03_120]→中隔縁束は卵円野ともよぶ。これは胸髄下部から腰髄の高さで、後索に入った後根線維内側群の下行枝が薄束の内側部、すなわち後正中中隔に接して集まってできた線維束である。後根を通過っして後索に親友する一次求心性感覚線維は、分岐して下行性に向かう側副枝を出す。下半身から脊髄に到達した感覚線維は薄束を形成し、その下行枝が中隔縁束をつくる。中隔縁束はコンマ束(束間束)と部位によって癒合することが認められ、また他方卵円野より腹側の正中域に中隔縁束と別の神経束として前中隔線維の存在も報告されている。
Triangular fasciculus; Olivospinal tract(三角束;オリーブ脊髄路)Fasciculus triangularis; Tractus olivospinalis さんかくそく;おりーぶせきずいろ
[A17_1_03_121]→ 延髄にある下オリーブ核から起こり、交叉性に脊髄側索を下行し、前角の運動ニューロンに接続する。 下オリーブ核は大脳皮質・線条体・赤核や脊髄(脊髄オリーブ路)からの入力を受け、骨格筋の活動を調節すると考えられている。(解剖学講義)
Rubro-olivary fasciculus(赤核オリーブ束)Fasciculus rubo-olivaris [A17_1_03_121_1]→
Triangulointercalated bundle(三角介在束;布施氏束)Fasciculus triangulointercalatus (Fuse) さんかくかいざいそく;ふせしそく [A17_1_03_121_1]→
Retroolivary fasciculus(オリーブ後束)Fasciculus retroolivaris おりーぶこうそく [A17_1_03_121_2]→
Ascending pathway(上行性伝導路) [A17_1_04_001]→
(皮膚感覚の伝導路) [A17_1_04_002]→
(脊髄神経による伝導路) [A17_1_04_003]→
(痛覚・温度覚の伝導路) [A17_1_04_004]→
(粗大触圧覚の伝導路) [A17_1_04_005]→
(精細触覚の伝導路) [A17_1_04_006]→
(脳神経による伝導路) [A17_1_04_007]→
(深部感覚の伝導路) [A17_1_04_008]→
(非意識型深部感覚の伝導路) [A17_1_04_009]→
(下半身の深部感覚の伝導路) [A17_1_04_010]→
(上半身の深部感覚の伝導路) [A17_1_04_011]→
(意識型深部感覚の伝導路) [A17_1_04_012]→
Optic pathways; Visual pathway; Visual tract(視覚路;視覚伝導路) しかくろ;しかくでんどうろ
[A17_1_04_013]→視覚伝導路は神経系の緒経路のなかで最も重要なものに属している。視覚伝導路に損傷を来す頻度も高いので、医師はその伝導路についての知識と理解を完全なものとそしておくべきである。 物体から眼球にすすんだ光は水晶体の屈折作用(逆転像形成をもたらす)を受けたのちに、眼球の神経層、すなわち網膜retinaにぶつかる。網膜には何種類かのニューロン(それには光受容性の桿体細胞、錐体細胞が含まれる)の層状配列がみられる。おのおのの目に耳側と鼻側の両視野が存在するが、水晶体の像逆転作用のために耳側視野は網膜の鼻側半に投射され、鼻側視野は網膜の耳側半に投射される。患者の部分的な視野欠如を記載するときには、網膜野ではなく視野で表現する方法が常に用いられる。眼球内の神経細胞から出た軸索は、視神経の中を後方に向かって走り、視神経交叉のところでは鼻側半網膜からの軸索が交叉し、耳側半網膜からの軸索(非交叉性)と合流して視索を形成する。視索のなかをさらに後方に走る軸索は、やがて外側膝状体に達し、ここで次のニューロンへの信号伝達がシナプスを介して行われる。外側膝状体から始まるニューロンの軸索は視放線を形成しつつ大脳表面に次第に近付き、後頭葉の視覚領皮質に達して終わる。視覚領皮質は後頭極よりはじまり、楔部と舌状回(二者の境が鳥距溝)にかけての広がりを示す。また、左右の眼球の左者情報が右の後頭葉皮質に伝えられる(左右眼球からの右視野上方は左の後頭葉皮質へつあえられる)点に十分注意された。水晶体の像逆転作用のために、上方視野上方が網膜の下半に、下方視野上方はその逆すなわち網膜の上半に、下方視野情報はその逆すなわち網膜の上半に、それぞれ到達することもわすれてはならない。このような上・下視野の区別は視覚伝導路の全体で維持されており、鳥距溝より上に位置している楔部が下方視野情報を、同溝よりも下に位置している舌状回が情報視野情報をそれぞれ受ける。最後に、黄斑と呼ばれる網膜の中心で最も鋭敏暗視覚の得られる部位からの神経信号は、後頭極に送られる。
Neuron of optic pathway(ニューロン(視覚路の)) にゅーろん(しかくろの)[A17_1_04_013_1]→
Auditory pathway(聴覚の伝導路) [A17_1_04_014]→
Taste pathways(味覚の伝導路)
[A17_1_04_015]→舌、喉頭蓋および喉頭部の味蕾にある化学受容細胞(図23)は、3種類の感覚ニューロンで支配されている(第5章-図8,10,11)。 感覚ニューロン。ニューロンの第1神経細胞体は膝神経節に位置する。それらは、顔面神経(VII)の中間神経を経由して脳に投射する。末梢の軸索突起は、顔面神経および鼓索神経、あるいは大錐体神経および耳神経節を経由して舌に投射する。両群の神経線維とも最終的には舌神経に合流し、舌尖近くを支配する。舌先端部に見出される茸状乳頭と葉状乳頭は、甘味、塩味そして酸味物質に対してとくに感受性が高いために、膝神経節の感覚受容細胞は、こうした3種類の基本味を感受する上でとくに重要である。 味感覚ニューロンの第2群は、舌咽神経下神経節に見出される。これらの神経は、中枢側では弧束核と結合し、末梢枝はは舌咽神経を経由して舌基部の味蕾と結合する。第3群のニューロンは、迷走神経下神経節に位置する。これらの神経は、迷走神経根を経由して脳幹の神経核に投射する。一方、末梢では、上喉頭神経を経由して喉頭蓋および喉頭に投射している。舌基部で、有郭乳頭中および喉頭蓋と喉頭中の味細胞は、とりわけ苦物質の発生に重要である。 中枢結合。 脳では、3群の味覚線維のすべてが弧束核の吻側部に投射し、味覚の二次中継ニューロンを活動させる。その後、これらのニューロンは、橋味覚野に投射する。そして、そこから視床、視床下部外側野および扁桃体に投射する。視床への投射は両側性に起こり、後内側腹側核(VPM)に終止する(図42)。VPM核は、視床における特殊中継核であり、感覚皮質で顔面の領域のすぐ下に位置する味覚領域に神経線維を送る。したがって、視床における味覚線維の結合は、意識にのぼる味覚の認知に関わっているらしい。味覚の投射が両側性であることは、味覚を全体として評価するのに役立っている。ただし、投射が拡がっていることによって、舌のどの部分で味を感じているかを限定する機能が比較的難しくなっている。一方、視床下部および扁桃体での味覚線維の結合は、主として味刺激に対する反射性、あるいは動機づけ反応、したがって、食物摂取の制御にかかわっているだろう(図62)。 味覚伝導路の核が唾液分泌に関係した自律神経核(上および下唾液核)と反射性の結合を脳幹で形成していることは、舌における食物刺激に対する味反応に伴った唾液分泌反射の解剖学的基礎を形成している。脳幹における他の自律神経系との結合は、“味覚性発汗”に関係している。これは、刺激性のあるいはコショウのきいた食物を摂取することに応じて起こる顔面および額部の発汗現象である。この反応は正常の反応ではあるが、発汗が異常に多い場合は病的と考えられている。 個々の味覚線維は、1種類の刺激にのみ応じるのではなく、程度の差はあるが、4種類全部の基本味に応じる。こうした味反応の多様性は、個々の味覚線維が複数の感覚受容細胞と複雑な結合をしていることから生じるらしい。図に示した味覚伝導のすべての段階でニューロンで、複数の味刺激に対しての反応がそれぞれ程度の差はあっても同じように生じる。さらに、視床VPM核での味覚感受性ニューロンは、機械的刺激には応じないが、舌の温熱刺激にも応じることが見出されている。個々の味の性質が、味覚伝導路を伝えられる混在した感覚情報からどのようなメカニズムによってひき出されるかはわかっていない。
(嗅覚の伝導路) [A17_1_04_016]→
Nervous pathways in spinal cord(脊髄の伝導路)Tractus medullae spinalis せきずいのでんどうろ
[A17_1_04_017]→白質中には上行性、下行性の主として有髄神経線維束が充満している。隣接する伝導路の境界は確然としているわけではない。後角先端と脊髄後縁との間の終帯は一見灰白質に見えるが、この部分は髄鞘の細い神経線維が占めている。

You are the th visitor to this page !(since 2000/1/1)
本日の番目です。昨日は
th visitor to this page !