Rauber-Kopsch解剖学
歴史的な偉大な解剖学書
Rauber Kopsch Band1. 05 


C.細胞の分化,組織および器官の形成Differenzierung der Zellen, Gewebe-und Organbildung
細胞の分化DifferenzierungすなわちSonderung(分別の意)は細胞分裂が持つ使命の1つであると思われる.細胞の分化はどれをもても,そのすべてが新個体の出発点である受精卵がひきつづいておこる分裂によって多数の細胞となって,これらの細胞が受精卵が受精卵から受けついだ材料に基づいておこっている.この分化の目的とするところは分業Arbeitsteilungである.分業は高等な生物には必要なことである.原性動植物はその体がわずか1個の細胞からなるので,この1個があらゆる役目をする.この場合でもある程度の分業があって,つまい核は原形質とは違った機能を持っている.なおまた原形質の内部にも分化がありうる.そこに筋原線維ができていることさえある.そして分化が必要である.そして文化の程度や形式がちがうのに従って,それぞれの生物が高低いろいろの段階に立っているわけである.
分化の量とその方向を知ろうとするならば,われわれはここであらゆる種類の細胞とそのつくったものまで数え立てねばならないのであろう.しかしその目的のためには,雑多な種類の細胞を無理の内容に分類してつまりいわゆる簡単な組織einfaches Gewebeの群に分けて述べるのがよいと思う.
組織とは,もともと同じ種類に属する細胞およびその細胞から生じたものの混合体である.
動物においてはそういう組織として4つが区別される.
1. 上皮細胞 2. 結合および支持組織 3. 筋組織 4. 神経組織
動物体の器官Organeはこの4つの組織のなかの1つあるいは2つ以上によってできている.器官という概念の形態学上および生理学上の定義は次のごとくである.
器官は1種あるいは2種以上の組織より成り,決まったか形と機能をもつものである.
その例として:1個の杯細胞は1つの組織に属するわずか1つの細胞からなる器官であり,爪は単一の組織に属する多数の細胞が集まってつくっている器官であり,腺はいろいろな種類の組織からできている器官である.
いくつかの器官によって機能的にいっそう高いつながりをもった単位が構成される.これが装置Apparatあるいは系(統)Systemである.例えば視覚装置Sehapparatは眼,眼筋,神経,血管およびその他の数多くの補助機関より成っている.
そして最後に動物体の全体は多数の器官や装置からできているわけである.
上述の関係からして,生物体の要素として細胞を述べたのに続いて,まず組織を列挙するには全くいろいろな方法がある.形態学的,生理学的,発生学的というそれぞれの立場がある.
これらの立場のどれを厳守してみても,組織の分類としては全般に妥当とするものとはならない.そこで最も広くおこなわれている分類がすぐ上に紹介されたものであって,これは単一の原則によって決定したのではなく,発生学と生理学と形態学の諸性質をあわせたものがその土台になっているのである.
Rauber, A., Die histologischen Systeme. Sitzber. Naturf. Ges. Leipzig, 10. Bd.,1883.-Haeckel, E., Ursprung und Entwicklung der tierische Gewebe. Jen. Z.,18. Bd.,1884.-Gaule, J., Oekus der Zellen; in: Beiträge z. Phys.,1887.-Peter, K., Die Gewebe im Unterricht. Anat. Anz., 68. Bd.,1930; Über anatomische und physiologische Eiheiten des Körpers. Anat. Anz., 71. Bd.,1931.-Patzelt, V., Zur Einteilung der Binde-und Stützgewebe. Acta anat. Vol. 9.1950.
IV.組織学Histologia, Gewebelehre
組織の分類にはいろいろな行き方があるが最も広くおこなわれいるのは次のものである.すなわち1. 上皮組織,2. 結合および支持組織,3. 筋組織,4. 神経組織に分けられるのである.
1.上皮組織Epithelgwebe
定義:上皮組織は細胞のみから成っていて,その細胞は接合質によりあるいは突起によりあるいはその両者によってたがいに結合しており,1層あるいは多層をなして連続し,体の内外にある自由表面を被っている.
これは細胞のみからなるので,あらゆる組織のなかで最も簡単なものであり,それゆえ細胞学のすぐあとにつづいて述べるのが好都合である.
一般的性状:上皮細胞は原形質と核より成り,はっきりした境界を示す細胞である.細胞膜はしばしば欠けて,原形質の周縁部が固くなって細胞膜の代わりをしていることが多い.大多数の上皮細胞は軟らかくて,従って周囲の圧関係に容易に適合できるのである.しかしまた正にその配列のために上皮細胞は事情によっては,かなり強い圧をたがいに及ぼしあうことができるのであって,そのために特に胎生期には形態発生の重要な諸現象を引き起こし得るのである.なお成体においても多くの上皮細胞団はその構成要素すなわち個々の脂肪がたがいにおよばす圧のもとにある.細胞の形が容易にそのことをしめしているが,その他の点からもこれは証明できるのである.
上皮組織をなす細胞の形が容易にそのことをしめしているが,その点からもこれは証明できるのである.
上皮組織をなす細胞の形の豊富さFormenreichtumははなはだ著しいものであり,大きさの差異も高度である.細胞体の外面および内部に多種多様な構造がみられることがある.
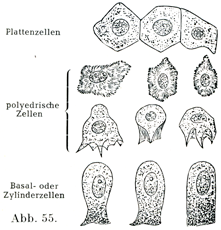
形状によって4種を分ける:扁平上皮Platten-Epithel(敷石上皮Pflaster-Epithelともいう),円柱上皮Zylinder-Epithel, 絨毛上皮Flimmer-Epithel, 移行上皮Übergangs-Epithelである.
扁平上皮の細胞は平たくて薄く,小鱗状の板をなし,その境界線が多くは不規則である.ただ網膜の色素上皮(色素層Stratum pigmenti)の細胞は規則正しい六角形をしている(図39).他の細胞,たとえば口腔上皮の表面の細胞は不規則な輪郭をしめす(図40).そのほかの扁平上皮細胞でも五角形あるいは六角形をするものがあって,その境界が直接的ではあるが,境界線の長さがまちまちである(図46).
円柱上皮細胞はいろいろの長さの稜柱の形をしている.それには(いわゆる)立法形の細胞kubische Zellen[「立法形」の細胞“kubische”Zellenという名前はよくない.さいころの形をした細胞を誰もみたことはあるまい.いわゆる立法形の細胞はその形から云えば五角形あるいは六角形の円柱状または稜柱状の細胞で丈の低いものである.(原著註)]からの眼の水晶体をなす長いひものような“水晶体線維”Linsenfasernまであらゆる形のものがある.楕円に近い形の核がその長軸を細胞の縦の方向に一致させて存在する.細胞の基底部は細い突起をだして,その下にある組織に鈎をかけたように付着している.
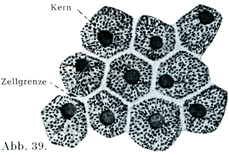
[図39]ヒトの網膜の色素上皮の細胞.
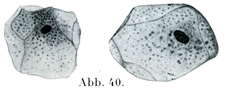
[図40]ヒトの口腔粘膜の扁平上皮細胞.×300.
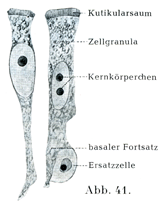
[図41]カエルの腸から取り出された円柱上皮細胞.×750.
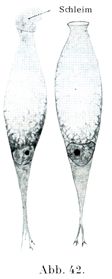
[図42]カエルの口蓋から取り出された杯細胞.×750.
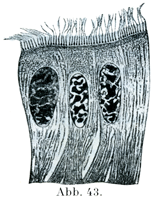

[図43~44]絨毛細胞.ヨーロッパの食用カタツムリHelixの腸上皮.(M. Heidenhain)
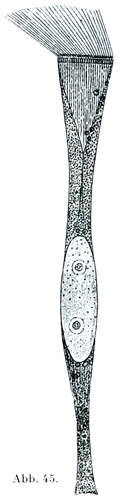
[図45]ドブシジミCyclasの絨毛上皮細胞.(W. Engelmann)
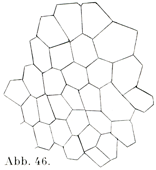
[図46]硬骨魚Perca fluviatilis(スズキ属)の小さい胚子の表皮を外方からみる.細胞の境をしめす.×200.
絨毛細胞Flimmer-oder Wimper-Zellenは円柱状または円錐状の細胞で,その自由面には多数の細かい毛が生えていて,その毛が細胞の生きているときは運動bewegen od. flimmernする.この絨毛は小皮縁Kutikularsaumという細胞体の縁の固い特別な1層付着している.そこで基底小体Basalkörperchenというものと結合している(図43~45).これからさらに細胞体の内部に細かい糸状物がのびて,これが集中しながら円錐状をなすので,絨毛根円錐Wimperwurzelkegelとよばれる.楕円に近い形の核が小皮縁からいくらか隔たったところにあり,その長軸は細胞じしんの縦軸と平行している.絨毛細胞の基底部は円錐状に細くなり,尖った突起をなしている.それが数本の細い糸状物に分かれていることがあって,これが上皮の下敷きをなすものに鈎で止めたように付着している.
一定の細胞でみると,絨毛の運動はいろいろと違ったぐあいにおこなわれるが,毛のなびく方向はいつも変わらないのである.小さい細胞で,しかも弱い拡大でみたのでは,絨毛細胞の特性はなかなか分からない.多くの動物のものを比較してしらべると,はじめて絨毛細胞の複雑な構造がいっそう明瞭にある.絨毛細胞は動物界の全体を通じて大きい役目をしていて,時としてはこれが唯一の運動器具をなしているのである(図45).
絨毛は単層の扁平上皮や高低のいろいろの円柱上皮にも存在することがある.後者は単層のばあいも銃創のばあいもある(図47).K. Peterの研究(Anat. Anz.,15. Bd.,1898,1899)によれば絨毛細胞の核をもっていない部分にも活発な運動がみられるのである.すなわち核は絨毛の動きには意味をもたない.またその毛だけを原形質の一部が付着しないようにとりだしても,やはりそれが運動するので,原形質もまたこの運動に直接の影響をもたない.絨毛運動をおこす中心はむしろ絨毛装置じしんのなかにあるのであって,基底小体のみがその責任を持つとおもわれる.この考え方と一致するのが精子Spermienの断片についての所見であって,中部Mittelstückとつづいている断片のみが運動をしめすのである.しかしなお基底小体と中心小体とが同じものであるかどうかが決定されていないし,また植物についての問題も容易に決まらない.(これについてはHeidenhain, Plasma und Zelle. Bd. I.,Jena,1907の287頁を参照のこと.)-v. Renyi (Zeitschr. Anat. Entwgesch.,81. Bd.,1926)は絨毛細胞の運動の中心に関する問題を新しい方法(生体染色,微小操作)を利用して研究した結果,“いままでの記載的および実験的な形態学において,絨毛細胞の運動の中心をいくらかでも確実さをもって決定することは,成功していない”という悲観的な結論に達したのである.
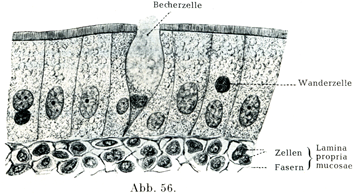
円柱上皮細胞や絨毛上皮細胞のそれぞれの集まりの中に,機能の上からは粘液細胞Schleimzellenとよばれ,形状の受けからは杯細胞Becherzellenとよばれる一種特別な細胞がある.その形は分泌物を含む量によって変わる.分泌物をほとんど有しないときは円柱細胞の観を円柱細胞の観を呈しているが,粘液が増していくると樽のような形になる.それも初めはまだ細長いが,だんだんと丸みをおびて膨れてくる.そうすると核は原形質の残りといっしょに細胞の基底部に移り,核の形が変わる.細胞の自由端のところが開いて粘液が密雲か綿雪のように膨れてその口から出て行く(図42, 47, 56, 57).
腸の杯細胞は円柱細胞が変わってこれになるのであって,後者の原形質内に粘液顆粒(図48)ができて前者が生ずる.粘液顆粒は始めはごく微細であるが,そのときすでに粘液反応を呈する.すなわち塩基性の色素に強く染まるのである.それより進めば顆粒が増大し,またたがいに癒合する.もっとも癒合は概して顆粒が細胞の外にでたうえで始めておこるのである.
他方また,ある腺の粘液細胞では粘液の前進をなす顆粒がみられることもある.その顆粒は最初のうちは粘液染色で染まらないので,あとになって粘液顆粒にかわる.Heidenhaln, Plasma und Zelle, Bd. I.-Osawa, G., Über Darmepithelien. Mitt. med. Fakultät Tokio. 9. Bd.,1911.
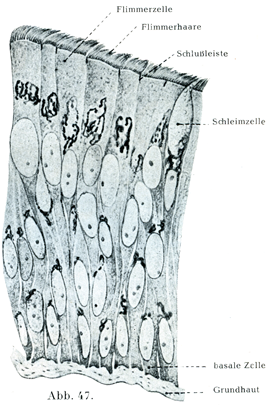
[図47]ヒトの気管の重層絨毛上皮.細胞内の内網装置をKopsch-Kolatschevの方法であらわしたもの.×1000. (Kopsch, Z. mikr.-anat. Forsch., 5. Bd.,1926)
移行上皮細胞は扁平上皮細胞とはなはだ近い関係にあるもので,尿の通路のみに存在する.すなわち腎盂,尿管,臍胱にある.移行上皮の最も表層にある細胞は,この上皮が被っている気管がひっぱられているときは,幅が広くて平らになり,その気管が縮むときは熱くなる.この細胞は自由面にそって小皮性の1層をもち,また細胞体の下面にはいくつかのへこみがあって,これにそれより深くにある西洋梨型あるいは紡錘状の細胞の頭が入り込んでいる(図49, 50).
多数の上皮細胞がたがいに結合して1つの上皮をなすこと,すなわち一と続きで隙間のない1層をなすことは接合質Kittsubstanz(細胞間物質Interzellularsubstanz),あるいは細胞と細胞とのあいだを通ずる細胞間橋Verbindungsbrucken (Interzellularbrucken),もしくはこの両者が同時に存在することによって実現している.
接合質の特別な一種であるところの閉鎖堤Schlußleistenは多数の上皮(それも諸所の粘膜の円柱上皮や移行上皮が主である)にみられるもので,これは表皮細胞の表面にむかった端のところを結合していて,表の方からみると閉鎖堤網Schlußleistennetzという網状の像をしめすのである.
細胞間橋は一つの細胞から他の細胞につづく原形質の突起と考える人があり,あるいは細胞膜の突起とする人もあり,またその両方だという人もある.その決定はむつかしい.もしもそれが原形質の突起だとすると,そしてこの考えは動物細胞は普通に細胞膜を欠くので確実性を有するのであるが,全身にわたる大きい細胞団の原形質が無数の突起でたがいにつづいているという重大なことになる.すでに胎児おいてこういう突起の存在が証せられる.すなわち早くから現われるもので,細胞相互のあいだを固く着けておくためのものである.他方ではまた細胞間隙Interzellularlückenという管が残っていて,そこを細胞間液interzellulare Flüssigkeit(上皮リンパEpithellymphe)という液が流動していて,これが上皮の栄養にはなはだ大きい意味をもつのである.遊走細胞もこの管を通って,上皮の表面に達する.
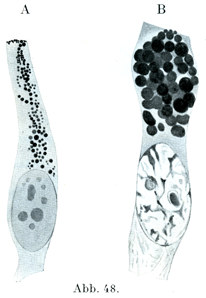
[図48]サンショウウォの腸上皮細胞における粘液形成.Aは小さい粘液顆粒をもつ細胞.Bは大きい粘液顆粒をもつ細胞.(Heidenhain, Plasma und Zelle. Bd. Iによる.)
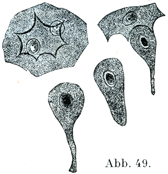
[図49]家兎の臍胱より得た移行上皮細胞.×300. 左上:表層の幅の広い平らな細胞で2つの核を有し,下面に鋭く突出した明瞭な縁とへこみをもっている.下:それより深い層に属する2つの西洋梨型の細胞.右上:西洋梨型の細胞が1個,表層の細胞がもつへこみのかなにはまっている.(Kleinに基づいてSchäferが描いたものよりとった.)
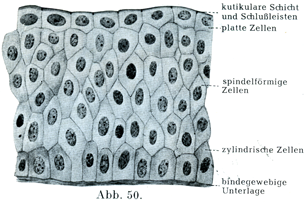
[図50]尿管の移行上皮 ヒトの尿管粘膜の横断図.×500.
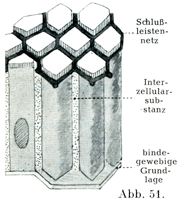
[図51]閉鎖堤網の模式図(Stöhr sen, による.)
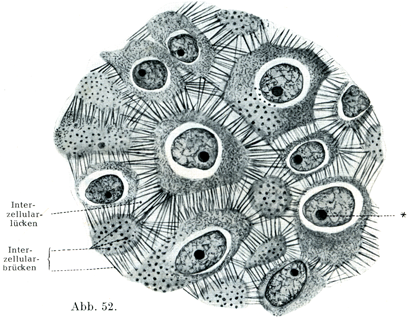
[図52]細胞間橋 ヒトの表皮の切片.×1000. 細胞間隙が広くなっている.そのために細胞間橋のランヴィエ小節Ranviersche Knötchenが存在しない.*は核小体.
上皮層形成:すでに述べたごとく上皮細胞が集まって,一とづづきの被い,すなわち上皮を形成している.重なり合っている細胞層の数によって
a)単層上皮einfaches (einschichtiges) Epithel
b)重層上皮geschichtetes (mehrschichtiges) Epithel
が区別される.
理論的に云えば,この二つの型は上述の4種の上皮細胞のいずれにも存在しうるはずであるが,しかし単層の移行上皮というものはない.だからわれわれは次の7種の上皮を見ることができる.
1. 単層扁平上皮einfaches(einshichtiges)PlatternEpithel:その例としては網膜の色素上皮,肺胞の呼吸上皮,精巣網の上皮,胸膜や腹膜の上皮(図39, 53).
2. 重層扁平上皮geschichtetes (mehrschichtiges) Plattern-Epithel:これに属するのはまず表皮,なお口腔,食道,声帯ヒダの自由縁,腟,角膜などの上皮である(図54, 55).
ここで注意するべきは,最も表面の細胞層のみが扁平な細胞からできていることである.最下の細胞層をなすものは円柱状であり,それに次いで多角形の細胞が集まっている.これは隣接する細胞から圧をうけて多角形となっている.表面の近くにある細胞は扁平であるが,最も表層にあるものだけが薄くて小鱗状である.細胞の形に応じて核の形および位置も変化する.核は下部の円柱細胞層では楕円に近い形であり,中くらいの層ではほぼ球形を呈し,上部の層では平たくなっている.
3. 単層円柱上皮einfaches Zylinder-Epithel:腸間では噴門から肛門までの上皮がそれであり,数多くの腺の導管や腺体(甲状腺,腎臓,前立腺,精嚢腺),脊髄の中心管,正看の上皮がこれがこれに属する(図56).
4. 重層円柱上皮geschichtetes Zylinder-Epithel:眼瞼結膜,大きい腺の導管の主幹,男の尿道,精巣上体管にこの種の上皮がある.
5. 単層絨毛上皮einfaches Flimmer-Epithel:気管支の細い枝,卵管,子宮,副鼻腔,精巣上体頭の管がこの種の上皮をもっている.
6. 重層絨毛上皮geschichtetes Flimmer-Epithel:呼吸道では鼻腔からはじまって気管支の細い枝まで,ただし声帯ヒダの自由縁をのぞく.そのほかに血管や鼻涙管がこの種の上皮で被われている(図57).
7. 重層移行上皮geschichtetes Übergangs-Epithel:これは腎盂,尿管,臍胱にある(図50).
上皮細胞の表面および内部にみられる分化äußereund innerre Differenzierungenははなはだしく多様である.それを全部述べることはくたびれ損の傾きがあるから,その大体を簡単に述べて,違った形のものが如何に豊富であるかを,ある程度わからせるとしよう.
上皮細胞の表面の分化のなかで,われわれはすでに細胞間橋と絨毛については述べたものである.動く付属物すなわち運動毛Kinocilienのほかに時として細胞の自由面に動かない毛すなわち不動毛Stereocilienがあるとされている.付属物のいま一つ別の型のものは精巣上体管などの細胞の方面にある細い毛の束であって,これは分泌物を導くはたらきをもっている(第II巻を参照のこと).
中心鞭毛装置Zentralgeißelapparatは精細な多くは短い1本の毛が細胞の自由面から外にむかって突出していて,鞭毛Geißelあるいは外糸Außenfadenとよばれ,その細胞の方面のすぐ下に多くは双心子Diplosomaの形で中心小体があり,なおこの中心小体から細胞の内部にむかって細い糸が集まって束をなしてすすんでいる.これらを合わせて中心鞭毛装置という.この鞭毛が動くかどうかはいままでまだ分かっていないが,精子細胞から最初に軸糸ができはじめる時と形態学的に似てはいる.しかしおそらくこの鞭毛は感覚器であろう.あるいは退化した構造物であろうか.というのはヤツメウナギの原腎の細胞にはかなり丈夫な鞭毛を1本ずつもつものがみられるのある.
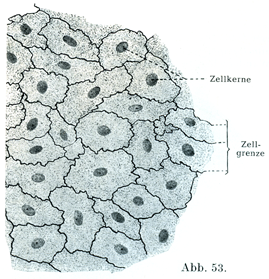
[図53]ヒトの大網における腹膜上皮 細胞の境界を銀によってあらわしてある.
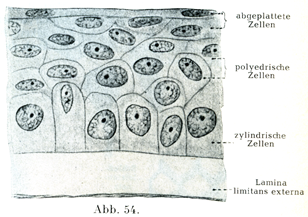
[図54]重層扁平上皮 ヒトの角膜上皮の切片.×500
特別な種類の突起が網膜の色素上皮の細胞にみられる.それは原形質の突起であって,色素顆粒をもっていて,この顆粒は細胞に光の当たるときとあたらないときで位置を変えるのである(図59).おのおのの細胞がそういう突起をたくさんにもっている.
また腸上皮細胞の自由面は独特な形態をしめしている.そこにあるStäbchensaum(小棒縁)[いわゆる小皮縁Kutikularsaumと同じものを指す.(小川鼎三)]あるいはPorensaum(小孔縁)の微細構造を正確につかむことは困難である.この縁がたくさんの縦の方向に平行したすじをもっていることは容易に分かる.しかしおそらくこれは細胞の原形質が指状の突起をたくさんに出して,それをとりまいて特別な小皮性の部分があるのであろう(図56).これについてはR. HeidenhainとOsawaの研究がある.
上皮細胞の外面の分化でいま一つ別の形のものはいわゆる刷子縁Bürstenbesatzである.Lieberkühnの腸腺,胃底腺,腎臓の迂曲した尿細管では分泌がおこなわえているあいだ,その明瞭さに多少の差はあるが細かい動かない毛あるいは小棒の形をしたものが細胞の自由単に衣服の辺飾りのように着ている.それが分泌のとき以外は消えてなくなるのである(Nussbaum, Arch. mikr. Anat., 27. Bd.,1886).
ここで感覚器の上衣細胞にみられるいろいろと特色のある分化について述べるべきであるが,それは感覚器の項にゆずることとする.
上皮細胞の内部の分化がまた実に多様である.その1つとして角膜および水晶体の上皮細胞はその原形質が全く明るくて,透明な性質をもっていて,光線がそこを通過するのに最もよく適している.そのちょうど反対が色素上皮細胞であって,その原形質は多数の小さい,不透明な小体を含んでいて光の通過をさまたげるのである.
いま一つやはり明るくて透明な,しかもはなはだ薄くひきのばされて核を失った上皮性のものが肺の呼吸上皮にある.ここにはその他に顆粒にとみ,核をもった小さい上皮細胞もある.
内部の分化として興味のある1つの型は上皮細胞の石灰化Verkalkungであって,これは正常な現象として,歯のエナメル質形成に起きる.長い棒の形をしたエナメル小柱のおのおのがそれぞれ1個の上皮細胞の石灰化した部分なのである.この石灰化上皮細胞Kalk-oder Titano-Epithelzellenに対して空気化上皮細胞Aëro-Epithelzellenというのがある.後者では空気が上皮細胞間の迷路中に侵入して,そおで栄養液を押しのけている.それは白髪の若干の場合,なお爪のしくみ得る場所がこの状態である.空気は細胞間隙から細胞じしんの中まで入り込んでいることがある.
全身にわたって大きい広がりをもっているのが角化Verhornungの現象である.角化は例えば毛の表面の毛小皮におけるごとく細胞の全体に完全におこっていることと,表皮の爪の角質層におけるごとく不完全なものとがある.角化の現象は典型的な場合には次のごとく進行する.ある細胞層にケラチンの前進をなすいわゆるケラトヒアリンKeatohyalinが粒状にあらわれて,これがそのとなりの層では液化しており,次いでこの液化したケラトヒアリンがさらにその向うにある角質化上皮細胞Kerato-Epithelzellenあるいは角質小鱗Hornschüppchenのケラチン膜の形成を引き起こすのである.
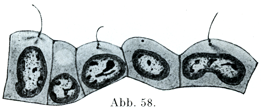
[図55]角膜の上皮細胞をばらばらにしたもの.×700.
[図56]円柱上皮(ヒトの回腸の切片)×1000.
上皮細胞の内部の分化としていま一つの別の種類は脂肪化上皮細胞Fett-oder Pio-Epithelzellenであって,その例は脂腺や乳腺の上皮にみられる.しかしこれらはわれわれが種々の上皮性の腺でみるところの細胞内部の変化についての万掌鏡ともいうべきもののほんのわずかな一部にすぎないのであって,それらの上皮性の腺については後に諸器官の項で述べることとする.
ここでは脊索Chorda dorsalisの組織について一言しておこう脊索は密に相接しあった細胞の集まりより成っていて,この細胞はやや進んだ段階では細胞膜をもち,細胞間橋(Studnicka)によってたがいに結合しているが,細胞間物質はごく少ししかなくて,そこに初めは膠原CollagenもコンドリンChondrinも含まれていない.しかしそれよりあとになると脊索組織の内部でそこそこに細胞壁が厚くなり,またその化学的性質がかわって,いわゆる脊索軟骨Chordaknorpelができる.それゆえ脊索の組織は学者によって結合組織に数えられ,あるいは上皮組織に数えられたりする(44頁を参照のこと).Schafferはそれが軟骨組織や結合組織に属するものではないと考えた.彼によればそれは支持物質の特殊な一型であって,宗族発生的に“軟骨組織の前進とみなされるものであり,軟骨様支持組織chordoides Stützgewebe”と呼ぶことができるのである(Anat. Anz., 37. Bd.,1910).-Studnicka, Z. Zellforsch.,13. Bd.,1931をも参照のこと.脊索の増殖した残物が成人にもなお存在している.
上皮細胞と神経線維
近年まで一般に信ぜられた学説は,神経の非常に細かい終末部が上皮細胞の間にある,すなわち上皮細胞間の液の流れている迷路部にあるというのであった.もっとも,それが細胞内にあると主張する声もなかったわけではない.近年のすぐれた研究方法によって初めて,ごく細かい神経線維が上皮細胞の内部に侵入して,その細胞体の中で,しばしば核に近いところで細かい終末網をもって終わることが確実に証明できるようになった.Boekeは“角膜のほとんどすべての細胞が神経をうける”と述べている.彼は扁平上皮を構成する普通の細胞および感覚細胞における細胞内神経終末を特に記載している.(図60~62).
Eggeling, H., Anat. Anz., 20. Bd.,1901.-Heidenhain, Sitzber. phys. med. Ges. Würzurg,1899.-Studnicka, Sitzber. Böhm. Ges. Wiss. Prag,1899.-Zimmermann, Arch. mikr. Anat., 52. Bd.,1898.
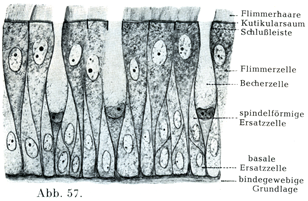
[図57]多列絨毛上皮mehrzeiliges Flimmer epithel. ヒトの鼻腔の呼吸部Regio respiratoria. [多列上皮mehrzeilings Epithelとは本質的には単層であるが,上皮細胞の丈が高低いろいろで,一部のみ表面に達する.従って核の並び方をみると多層のごとくおもわれるものをいう.(小川鼎三)]
[図58]中心鞭毛装置 ヒトの精巣網の上皮細胞.(Alverdes, Z. mikr.-anat. Forsch.11. Bd.,1927より.)

[図59]2個の色素上皮細胞 ヒトの網膜,細胞の側面からみる.細胞の下部は色素を欠き,上方に向かって長い絨毛状の突起ができている.核は示されていない.(M. Schultzeによる.)
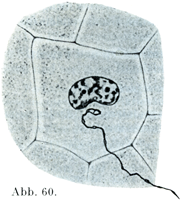


[図60~62]上皮細胞における細胞内神経終末 (Boeke, Zeitschr. mikr.-anat. Forsch. 2. Bd.,1925より.)図60. チゴハヤブサBaumfalkenの角膜上皮の細胞.図61. ハリネズミの輪郭乳頭の扁平上皮細胞.図62. ハリネズミの味蕾の内部細胞(感覚細胞).
2.結合および支持組織Bindesubstanzgewebe
定義:結合および支持組織は細胞間物質(あるいいは原線維間物質),細胞および線維より成っている.
この組織は中胚葉に由来するもので,動物体の支持器官をなし,血液およびリンパをつくる.
このグループに属する組織はいろいろあって,互いのあいだで重要な多くの差異を示すが,その起原が一致すること,他のものに変形する性質があること,比較解剖学的に特徴があること,およびまた種としてその機能の点で,完全な1群にまとまるのである.これらの組織の大部分をまとめて特別な1群となして,これに結合物質Bindesubstanzという名前をつけたのはReichert 1845が初めてである.
上に述べた3つの成分はこの組織群に属するいずれの種類の必ず存在するが,その中で最も本質的なものは細胞であって,これから他の2成分ができるのである.つまり細胞が細胞間物質および原線維をつくる.-早い時期の胎児では始めに結合組織Bindegewebあるいは間葉(間充組織ともいう)Mesenchymとして細胞が集まって網状を呈していく(図63),この細胞が突起をだしてたがいにつづいている.間葉の細胞は結合および支持組織のいろいろの種類ができてくる源なのである.
細胞間物質Interzellularsubstanzは細胞や線維の間にある無構造ののもので,従ってその名前のごとく細胞間のみでなく,また原線維間物質Interfibrillarsubstanzなのである.
Schaffer(Anat. Anz.,19. Bd.,1901)はこの無構造の中間物質を接合質Kittsubstanzとよんだ.v. Korff: Merkkei. u. Bonnet, Ergebnisse,17. Bd.,1909をも参照のこと.R. Virchow, Kollikerやその他の学者によればこの物質は結合組織細胞から分泌されて生ずる.この“分泌説”Sekretionslehreに対して比較的新しく別の学説が登場している.それは細胞間物質が細胞の外形質Exoplasmaの変形によって生ずるという“外形質説”Exoplasmalehreである.それにあとで細胞からの分泌物が加わるというのである.Heidenhain, Plasma und Zelle. Jena,1907-1911.-Stundicka, Sitzber. Bohm. Ges. Wiss. Prag,1907, Anat. Znz.1907, Anat. Anz., 38. Bd.,1911. Z. Zellforsch., 4. Bd.,1926.
基質Grundsubstanzという名称は原線維と原線維間物質ないし細胞間物質をみな含めたものに用いられる.結合組織の線維の由来については後述39頁を参照のこと.
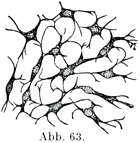
[図63]胎児の結合組織 ×400(Gegenraurによる.)
結合および支持組織の形状はあまりにも多岐にわたっているので,その分類ということが特に必要である.Waldeyerは次のように分類して,学生にこのむつかしい部分をできるだけ楽に会得させようとした.彼がいうごとく:
この組織は細胞間物質と細胞と線維とから成っている.その第1群では線維が他の2成分よりもずっとめだっている.それに属するのは
1. 普通の疎性結合組織gewöhnliches lockeres Bindegewebe.
内筋周膜Perimysium internum, 神経内膜Endoneurium, 腺の間やその内部,皮下組織などがそえである.
2. 定形(あるいは強靱)結合組織geformtes (straffes) Bindegewebe.
腱,筋膜,腱膜,眼球外膜Tunica exerna oculi, 薄膜Albugineaなどがこれに属する.
3. 弾性組織elastiches Gewebe.
平滑筋線維の腱,椎弓間靱帯Ligg. inerarcualia,肋間靱帯Ligg. intercotalis,血管壁の中などである.
第II群は細胞間物質が他の2成分よりも著しくみえるもので,これに属するのは
4. 軟骨組織Knorpel-Gewebe.
a)硝子軟骨
肋軟骨あるいは関節軟骨,また喉頭,器官,鼻そのほか数カ所にある.
b)線維軟骨
椎間円板,関節半月と関節円板,大腿骨頭靱帯,関節唇,なお所により腱や腱鞘のなかにある.
c)弾性軟骨(あるいは網状軟骨).
耳,喉頭蓋,そのほか喉頭の披裂軟骨の一部,小角軟骨,楔状軟骨がこれである.
5. 骨組織Knochengewebe.
骨格をなしている骨やいくつかの感覚器にある.また筋系統とともに近い関係にある種子骨や腱の骨化もこれに属し,歯ではセメント質がこれである.なお喉頭軟骨および肋軟骨が年齢が進むにつれ骨化することもここで指摘しておく.
6. ゾウゲ質組織Zahnbeingewebe.
これは歯にだけある.
第III群は細胞そのものが最もめだってみえる場合である.
7. 脂肪組織Fettgewebe.
体じゅうどこにもあるが,とくに皮下組織,腸間膜,腎臓の脂肪嚢でよく発達している.
8. リンパ性組織(あるいは細網組織)lymphoides (cytogenes, adenoides, retikuläres) Gewebe. リンパ節,内分泌腺,骨髄,胸腺などにある.
9. 色素組織pigmentiertes Gewebe.
眼球中膜tunica media oculiがその例.
10. 内皮組織Endothelgewebe.
血管やリンパ管を被い,またリンパ腔Lymphräumeを被っている.
11. 胎児性結合疎域embryonales Bindegewebe.
胚子embryoや胎児Fetusの体にある.
1. 普通の疎性組織結合は細胞間物質(これは同時に原線維間物質でもあるが),膠原線維,弾性線維,格子線維,固定結合組織細胞,遊走細胞,顆粒をもつ細胞(これにはEhrlichの肥満細胞Mastzellen, Waldeyerの形質細胞Plasmazellen, Ranviewの断裂細胞Clasmatocytenがある)からできている.それになお少数の脂肪細胞や色素顆粒をもつ結合組織細胞が加わっている(図64).
線維は一見したところ不規則に入り乱れており,いろいろな方向に走ってたがいに交わっている.外観により,また物理的および化学的性質の違いから,a)膠原線維collagene (leimgebende) Fasern, b)格子線維Gitterfasern, c)弾性線維elastische Fasernを区別する.
膠原原線維collagene Bindegewebsfirillenは非常に細いもので,その長さはしばしばはなはだ大であり,色は淡白で,均質にみえて,条らしい模様を示さない(近年,電子顕微鏡の利用によって膠原原線維に横縞のあることが確証された.(小川鼎三)).これはコラゲンCollagenという物質よりなっていて,煮ると膠Leim (Glutin)ができる.この線維が孤立していることもあるが,またいろいろな太さの束に合している.少量の原線維間物質がその間をくつつけ合わせている.
こういう結合組織束Bindegewebsbündelは繊細な縦の方向の筋を示し,かなりの丈夫さをもっている.そして曲がりやすいが,弾性は少ない.
結合組織束は同じ太さのままで長くつづいて軽い波状をなして走ることもあり,また同じ種類の他の線維束と合して,いっそう太い二次束や三次束をなすこともある.
試薬に対する関係:石灰水Kalkwasser,重土水Barytwasser,過マンガン酸カリ,ピクリン酸などを用いることによって線維束はその個々の原線維を分けられる.これらの化学物質が原線維間物質を溶かすからである.薄い酸たとえば酢酸を加えると,線維束は膨化して,膠のように透明になる.膠原線維は胃酸に溶けるが,トリプシンには溶けない,そしてすでに述べたように煮ると膠ができる.年齢による変化:若い人の膠原線維は上に述べたいろいろのものを作用させたときにいっそう弱い抵抗を示す.年齢が増すとともにますます固くなり,抵抗が強くなる.
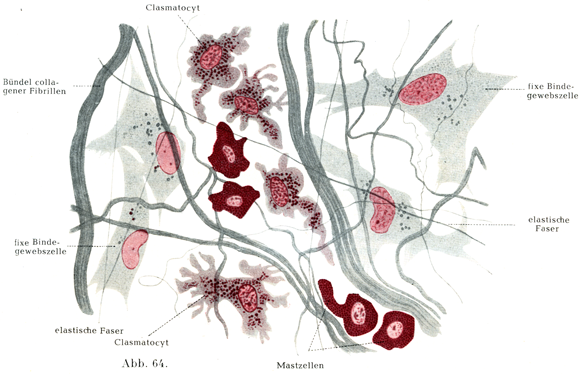
[図64]疎性結合組織 材料はネコ,中性赤による超生体染色supravitale Färbung mit Neutralrot. (Maximow)
格子線維は多くははなはだ細くて,たがいにつづいて格子状をなし,また膠原原線維の端とつづいている.薄い酸に対しては膠原線維よりも抵抗がいっそう強い.トリプシンによって消化されない.しかし煮ても膠を生じない.
格子線維は腺の終末部のまわりや毛細管の内皮管のまわりをとりまく基礎膜を形成している.銀液で処理するとこの線維が特別によく現われるので,そのために銀好性線維argrohile Fasernとよばれる.細網組織や脾臓の静脈洞をとりまく輪状線維は格子線維に属するのである.電子顕微鏡の示すところによると,この格子線維はいくつかのさらに細い原線維の集まりである(v. HerrathとDettmer, Z. wiss. Mikr. 60. Bd.,1951).
弾性線維elastische Fasernは多くは円い切り口で,時としてはまた帯のようである.これははなはだ弾性にとんでいる.その太さは1µの何分の1から9µまでいろいろと異なる.たがいに連なって網をなし,また線維の輪郭がはっきりとして暗くみえ,光を屈折する力が大きいので強い輝きをもっている.この線維が(標本のなかで)千切れると,その強い弾性のために切れ端のところが縮んで,特徴のあるうねった像をあらわす(図64).
試薬に対する関係: 弾性線維は酸およびアルカリに対してはなはだ抵抗が強い.しかしトリプシンによって徐々に全く溶けてしまう.水の中では60時間煮ても溶けない.圧を加えて30時間130°で煮ると弾性組織は特別な化学性状をもつ褐色の塊に変わる.
固定結合組織細胞fixe Bindegewebszellenはまた線維細胞fibrocytenあるいは線維芽細胞Fibroblastenともよばえ,その細胞体は扁平で,平らにひきのばされて尖った輪郭を示し,平板状あるいは翼状の突起をもってその突起が他の細胞の突起とつづいている.原形質は網状の配置を示して,小さい顆粒がわずかに含まれている.突起の縁のところは極端に細かくて薄いので,切片標本ではこの細胞の輪郭がほとんど決定できない.中心小体ももちろん存在している.内網装置はPfuhl (Z. Anato. Entw.,99. Bd. )によりはなはだ明瞭に染めだされた.
核はかなり大きくて,扁平で楕円に近い形をなし,数多くの細かい淡色のクロマチン顆粒と明瞭な核膜と1個あるいは数個の核小体をもっている.
これは結合組織の線維をつくるもので,高度に分化した細胞である.Maximowはその増殖によって同じ種類の細胞のみができるといい,Verattiは条件によっては之から形質細胞が生ずるという.またv. Möllendorff (Münch. med. Wochenschr.,1926, Z. Zellforsch. .,12. Bd.,1931)によると断裂細胞そのほかの細胞も之からできるという.色素を多量にあたえたときに限って,固定結合組織細胞はその色素の一部を自らの体内にとりこむが,それも細かい顆粒の形でみられるのみである.
遊走細胞Wandcrellen (Haemocyten) (図65)は形状および外観がいろいろである.原形質をわずかしかもたない小さい球形の細胞で,その円い核が血液のリンパ球の核のように濃密な作りを示しているものがあるかとおもえば,それより図体が大きくて,細胞体が良く発達しているのがある.おそらく上述の小さい遊走細胞がさらに発達してこれになったとおもわれるのである.遊走細胞はアメーバ様の運動を示し,その核は球形,馬蹄形あるいは分葉形である.
顆粒をもつ細胞granulierte Zellenとしては次の3型がある:
1.肥満細胞Mastzellen (ehrlich), 2. 形質細胞Plasmazellen (Waldeyer),3. 断悦細胞Clasmatocyten (Ranview).一部の学者によるとこれらの細胞は白血球に由来するという.そしてこれらに共通なことは細胞体が細大の差こそあるがいずれも顆粒をかなり沢山にもっていることである.
肥満細胞(図64)はずんぐりした大きい細胞である.水に溶ける粗大な小球状の顆粒が細胞体をぎっしり充たしていて,そのために核がしばしば全くかくされている.この顆粒は塩基性アニリン色素に染まる(metachromatisch, 染色に使う色素と違った色調で染まる).
この種の細胞は血管のそばや上皮層の近く,ならびに若い疎性結合組織のなかにみられる.動物が飢えた状態になっても,肥満細胞は存在しており,その特色を保っている.だから動物の全身的な栄養状態や局所的に高まった栄養状態とこの細胞は無関係である.それゆえ,肥満細胞という名前はむしろ細胞じしんが顆粒に富んでふくれていることにあてはまる.この細胞はヘパリンおよび若干の段階のヴイタミンを有している(Hirt, Verh. anat. Ges.1938).
形質細胞の形はいろいろで,円いこと,卵円形のこと,紡錘状のこともあり,数個の突起をもつこともある.原形質は塩基性アニリン色素で暗く染まるが,明瞭な顆粒構造を示さない.この細胞は主として血管の周囲に存在する.核は球形で,少数の大きいクロマチン小塊が核膜の内面に付着している(車輪核Radkern) (図65).
断裂細胞は組織球Histiocyten (Goldmann 1909),または大食細胞Makrophagen (Metschnikoff 1892),または休止遊走細胞ruhende Wanderzellen oder Polyblasten (Maximow 1906),または断裂分泌細胞rhagiocrine Zellen (Renaut 1907),または外膜細胞Adventitiazellen (Marchand 1898)とよばれるものと同一の細胞である.その細胞体は大きくて,多くの場合紡錘状を呈し,細胞体が広がっていて,顆粒に富み,また多数の液胞によって貫かれている.アメーバ様の運動をする.断裂脂肪という名前のおこりはその細胞体が部分的に切れて,その離れた部分は直ちにこわれてしまうが,核をもつ細胞体の主部は再生するという特性にある.Ranvierはこれを一種の分泌現象とみなしたものである(図64, 65)
この細胞は色素を豊富に,しかも粗大な塊の形で貯える.また明らかに“正常の結合組織内で物質代謝についての重大な役目を果たしている”(Pfuhl 1933).しかし寿命の短い細胞であって,これに反して線維細胞は寿命が長い.(Pfuhl, Z. mikr.-anat. Forsch., 31. Bd.,1932).
疎性結合組織にはなお脂肪細胞や色素をもつ結合組織細胞もまた,存在しうるのである.これらの細胞の構造についてはあとに詳述する.
組織発生:組織内白血球Gewebsleukocyten, 休止遊走細胞,組織球,大食細胞,外膜細胞はv. Möllendorff (Z. Zellforsch., 3. Bd.,1926)によれば線維細胞が突起をもってたがいに連なっている結合状態からその一部が離れてでて,その形態と機能を変じたものである(しかしこの点についてはMaximow, Leopoldina, 4. Bd.,1929およびBrodersen, Z. mikr.-anat. Forsch.,14. Bd.,1928を比較参照すること).
膠原線維は固定結合組織細胞からできる.それゆえこの細胞が線維芽細胞Inoblasten oder fibrobblastenと呼ばれるのである.線維がどこでできるかについて2説が対立している.Flemming, Spuler, Retterer, Maximowによるとそれは線維芽細胞の内部に生ずる.他方,Kölliker, とMerkelによればそれは細胞と細胞の間で,はじめ均質みえる細胞間物質の中にできてくるのである.前の説では,細かい線維芽細胞体の周辺部の明るくて均質にみえるそう,すなわち外形質Exoplasmaの内部に現われる.それも細胞の並んでいる列の全体をこえて一と続きのものが同時にできる.次いでこの線維が生みの親である細胞から分離して,そしておそらくは細胞から独立してなお成長することができるのである.もしもある定まった場所に線維芽細胞の大きい集団があって,その各々の細胞が高度の活動をなすならば,そこから線維に富んだ力強い結合物質の器官が生ずる.その反対に,少数の線維芽細胞が散在していて,それがわずかな活動をするときは,繊細な膜あるいは薄い繊弱な止め紐しかできないのである.
原線維の形成が終わったあとでは,細胞はできあがった結合組織性の器官の形状や組成に応じて,いろいろと変わった形をとらなければならないのであって,その形は周囲から細胞が押しつけられて生ずるのである.
かくしてわれわれは紡錘状や星形をしたもの,そして網状にたがいに合している細胞をみるし,また上皮のように細胞が沢山ならんで(上皮様細胞epitheloide Zellen),原線維の一次束の中に切りこんでいるのもみられる.この上皮様細胞が数個の束に同時に接するばあいには,非常に奇妙な形をとって,翼細胞Flügelzellenや水車の水掻き細胞Schaufelradzellenとよばれることがある.そのときには原形質は核の近くにだけ残っていて,細胞体の端っこのところは明るい板になっている.
上皮様細胞で囲まれている1本の原線維束に酢酸を加えると,その原線維の塊が著しく膨れるが,その膨れに対して上皮様細胞の包みが多少とも妨害をおこすのである.つまり所々で外に向かって膨れるが,その間のところがくびれている.その絞れは細胞の比較的つよい突起のある場所に当たっている(図67).そしてこの突起がいわゆる巻酪線維umspinnende Fasern (umsponnene Fasernというのがいっそう良い)である.Watzka (Z. mikr.-anat. Forsch., 40. Bd.,1936)によるところの絞れは膠原線維束のまわりを輪状にとりまいている格子線維によるのであって,薄い酸に対して格子線維が膠原線維よりもいっそう抵抗が強いので上述のことがおこるのである.
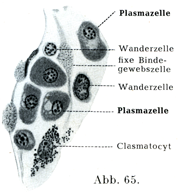
[図65]形質細胞 家兎の網Omentumより.(Maximow)
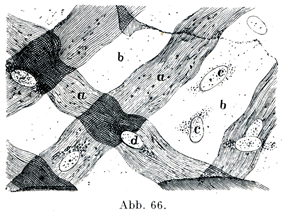
[図66]結合組織の薄い膜 皮下細胞に富む組織から,(A. KeyおよびG. Retziusによる.)約×750. a, a原線維の集まった帯,b, bこの膜の細胞層がたがいに合する場所,c, cその細胞の核,dこの膜の細胞核が原線維の帯の上にのっている.
2.定形結合組織すなわち強靱結合組織ではそのほとんど全部を占めて膠原原線維と特別な形の細胞,ならびに少量の細胞間ないし原線維間物質がある.原線維はたがいに平行して走り,わずかな量の原線維間物質のよってたがいに連ねられて,太い線維束をなしている.腱ではこの線維束がたがいに平行に配置され,筋膜や角膜ではそのかなり大きい集まりがたがいに交叉しており,真皮や眼の強膜では比較的小さい集まりがいろいろの方向に交錯している.細胞としてはほとんど結合固定結合組織細胞のみであるが,,これがそれぞれの器官に特有な,そして特別な形をしており,且つ規則正しい配置をしている(例えば腱の翼細胞,角膜の角膜小体).翼細胞Flügelzellen(Ranvier)は稜柱状の細胞であって,平たくひろがった3つあるいはそれ以上の数の突起をもっている.この突起はいろいろと異なる角度で細胞体から出ていて,隣接する線維束を囲む形をしている(図69).この種の細胞を側面からみると,視野の面にある翼状突起は薄い板としてみえるが,観察者の方に向かってのびているか,あるいはその反対の方向に伸びている突起は細胞の上を越えて通るくらい線としてみえるのである(図70).核は楕円に近い形である.角膜の細胞(角膜小体)は1つの面上にひろがった扁平な細胞である.
3.弾性組織は大部分が弾性線維ないし弾性網あるいは弾性板と少量の細胞間ないし原線維間物質,膠原線維および細胞とからなる.この組織は特別な弾性器官をつくっている.このような器官では弾性線維が結合してやはり網の形をしているが,その線維が疎性結合組織などの含む弾性線維よりはるかに太い.その例として項中隔の横断を示しておこう(図71).ここでは器官の軸にそって縦走する太い弾性線維の群が膠原線維と細胞間物質と細胞の3要素からなる疎性結合組織によってまとめられいる.
血管の平滑筋線維のあいだおよびいわゆる弾性軟骨では弾性組織が上述のものとは違った形で存在する.これらでは密な弾性網が融合することによって広い板や膜ができている.これはHenleいらい有窓膜gefenstere Häuteと呼ばれている(図72).

[図67]巻酪線維umsponnene Faserヒトのクモ膜,酢酸をもって処理したもの.(Toldt)
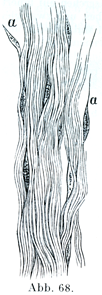
[図68]腱の膠原原線維Bindegewebsfibrillen (Rollet)約×500. a, a紡錘状の細胞とみえるもの.
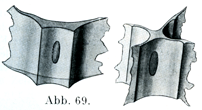
[図69]腱細胞の立体図plastische Darstellung. (Tourneux)
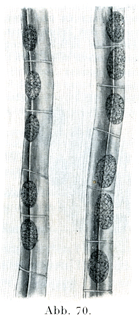
[図70]腱細胞の列 ネズミの尾Rattenschwanzより.
4.軟骨組織の特色は基質が方に存在して,その基質は透明であり,一見均質のごとくみえ,これを煮るとコンドリンChondrinが生ずることである.軟骨の基質は原線維間物質,膠原線維ないし弾性線維からなり,なお特徴ある軟骨細胞をその中にもっている.
いま述べた成分のいずれが他に優って存在しているかによって3つの主な群を分ける.
a)硝子軟骨hyaliner Knorpel, b)線維軟骨Faserknorpel(また結合組織軟骨Bindegegewebsknorpelともいう), c)網状軟骨Netzknorpel(あるいは弾性軟骨elastischer Knorpel),なお付け足りとしてd)細胞軟骨Zeenknorpelおよびe)石灰化軟骨verkalkter Knorpelを数えることができる.
石灰化軟骨を除けば,軟骨はすべて丈夫で弾性があり,色は青みをおびるもの,乳白色のもの,あるいは黄色のものがあり,特に胎児の体で,しかしまた成人の体においてもはなはだ重要な役目をなしている.その役目は軟骨の弾性や固さ,ならびにその表面が平滑であることによって達せられている.
a)硝子軟骨はガラスのように透明な一見均質と思える基質をもっている.その中に大きい円形あるいは多角形の,そして繊弱な感じのする軟骨細胞が2個あるいはそれ以上の数のものが集まって存在する.その原形質は細かい顆粒を示し,脂肪滴やグリコーゲンをもっていることがある.核は球形で繊細な核材を示す.脂肪体はいろいろの試薬にはなはだ敏感である.処理が不適当なときには細胞体が縮んで,とげをもつ小塊となる.生体内あるいは個体が死んでもなお組織が生きている場合には軟骨細胞はそれを容れるための基質内の滑らかな壁を持つ部屋,すなわち軟骨小腔Knorpelhöhleを全く充たしているのである.軟骨小腔の内面に接するところは他の基質の部分とは化学的にやや異なっていて,抵抗もいっそうつよいのである.ここを軟骨小嚢Knorpelkapselという.
1つの軟骨小嚢に包まれて2個の細胞をみることが稀でない.またそういう2個の細胞のあいだに薄い硝子様の隔壁のある場合もある.細胞が急激に分裂して,そのとき隔壁の形成がおこらないときは,細胞のかなり大きい群が1つの軟骨小嚢で囲まれる(図73, 74).
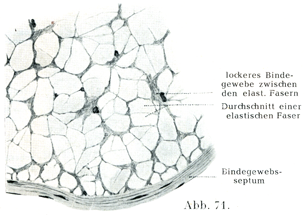
[図71]ウシの項中隔Lig. nuchae, Nackenbandの横断.
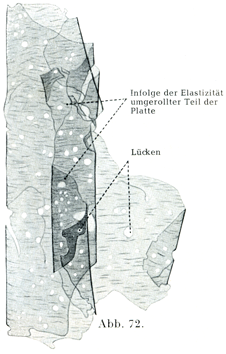
[図72]弾性板 ヒトの大腿動脈より得たもの.×75
意見したところ,どこも同じ性質であるかと思える基質が,ヘマトキシリン,ビスマルク褐,チオニンなどの色素に染まる傾向をつよくもっている.基質は大部分が細かい原線維より成り,これが原線維間物質によってまとめられている.過マンガン酸カリや食塩水をはたらかせたり,トリプシンで消化させたりすると,基質の組成がよくわかる.原線維は集まって束をなし,この束が多くの層をなして交叉もしくは交錯している.原線維亜また,軟骨小嚢の物質をも貫いているようである.
比較的年配のヒトの肋軟骨の内部,また関節軟骨や骨の軟骨結合のところに,老人性変化として基質が粗な線維の形をなして避ける現象が起こる.その個所は肉眼的にアスベスト様の観を呈するので,石綿様変性asbestartige Degenerationとよばれる(図74).また年配のヒトの軟骨細胞では脂肪の含有量が増す.
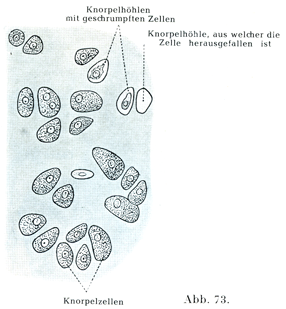
[図73]硝子軟骨 カエルの新鮮な材料で関節軟骨より切片をつくったもの.図の中央部にある明るい隙間で,その中に小さい顆粒を示すものは標本作製のときにこわされた細胞の残りを含む軟骨小腔の一部を現わしている.(Schiefeerdecker und Kossei, Gewebelehreより.)
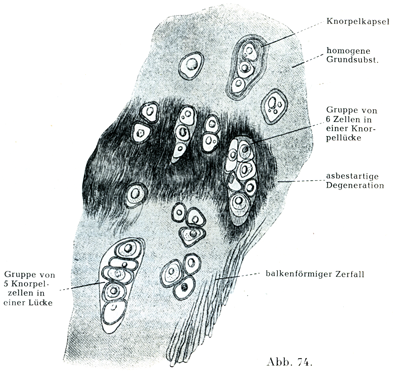
[図74]比較的年配の男の肋軟骨.横断.(Freyによる.)
軟骨は初めはその内部を貫く血管を全く有しない.そのはたらきをもっているのは軟骨の表面を被っている結合組織膜すなわち軟骨膜Perichondriumの血管である.しかしその後に軟骨が大きくなったときには軟骨膜から血管が軟骨の内部に侵入する.そして成人の軟骨ではふたたび血管がみられなくなるのが普通である.しかし奥深く存在している軟骨細胞も栄養を受けなければならない.そうすると基質の内部に特別な液細管Saftbahnenがたぶん存在するのであろうか?いままでこの点はずいぶん研究されたが,まだ確かりしたことがさっぱりわかっていない.もっとも染色の仕方によると,基質の中にはなはだ立派な網の形をした構造があらわれて,その網の形をした構造があらわれて,その網の結び目にあたって軟骨小嚢がある.基質内の液の流れに関係を持つ構造がこれであるまいかと考えるのも無理からぬことである(Schiefferdecker).
軟骨の石灰化Verkalkungは次のごとくにしておこる.まず軟骨小嚢のところに炭酸石灰が小さい顆粒の形で沈着するので,細胞はこの石灰化した軟骨小嚢によって全く包まれる有様となる.ついで石灰沈着は基質の残りの部分に及んでいくのである.
喉頭や器官の軟骨では真の骨形成Knochenbildungが軟骨組織を押しの小手,しばしばおこるのであるが,これはChievitzによればすでに20才ないし22才ではじまるのが普通である.これはほとんど正常の現象とみなされる.
軟骨の発生Entwicklung des Knorpelsは次のようにしておこる.若い未分化の結合組織細胞が大きくなって,組織細胞の原線維をつくる.この原性形成がすむと,線維芽細胞が前軟骨細胞Vorknorpelzellenと軟骨細胞に変化する.
これで軟骨形成の最初の原線維星の時期が達せられたわけである.その後に原線維が均質な接合質の沈着によって目に見えない状態“Maskiert”になる.かくして硝子軟骨の基質ができたのである.線維軟骨と弾性軟骨では線維が目に見える状態にある.v. Korff, Arch. mikr. Anat.,84. Bd.,1914.
しかしまた,軟骨の発生には間接的indirektなでき方がある.つまりすでにでき上がった他の組織すなわち軟骨膜あるいは小細胞性kleinzelligの結合組織から出発して軟骨が生ずるのである.
一とたび生じた軟骨がさらにひきつづいて成長するのは一部は間接的interstitiell, 一部は付加的appositionellである.付加的というのは軟骨膜の深部の層が軟骨組織に変化して,かくして軟骨の厚さがますことである.
軟骨の物質欠損Substanzverlusteの補充は温血動物では徐々におこるのみで,欠損が大きいとその補充は不完全にしかおこらない.そのさい再生現象は軟骨膜から出発する.
硝子軟骨は上に述べた場所のほかに,またすべての線維軟骨結合や軟骨結合において骨に密接したところにある.なおまたそのほか骨の数カ所にみられる(長腓骨筋腱溝,翼突鈎溝,小坐骨切痕,アキレス腱の踵骨終止部).一石灰化した軟骨は人間では特に関節軟骨にみられる.それは硝子性にとどまっている部分とそれにすぐ続く骨との間のところにある.
b)線維軟骨はその量の大部分をなすのが膠原原線維の束であってこれらの原線維は少量の原線維間物質によってたがいに結合しており,またいろいろの方向に走ってたがいに交叉している.その間に孤立して,あるいは群をなして軟骨細胞が存在し,これらの細胞のおのおののが少量の軟骨基質によって囲まれている(図75).
c)弾性軟骨(図76)は硝子軟骨の一種で,その基質内に弾性線維および弾性板の密な網を有するものである.軟骨の表面に向かってこの弾性組織はだんだん少なくなる.そしてこれは軟骨膜の弾性線維と直接に続いている.

[図75]線維軟骨 ヒトの椎間円板より.細胞のまわりの同心性の線は基質が順々にときを追うて生じたことをあらわしている.
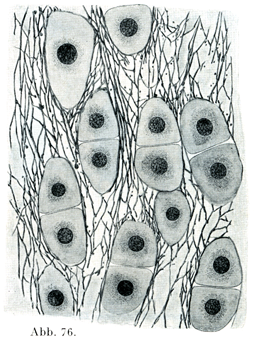
[図76]弾性軟骨 ヒトの耳より.×1000.
細胞軟骨Zellknorpelは成人には存在しないもので,軟骨細胞と軟骨小嚢,ならびにごくわずかの基質からなっている.脊椎動物の発生初期にみられる多くの軟骨,いろいろの哺乳動物の外耳の軟骨,また多くの魚類の鰓の小突起中にある軟骨の一部などがこの細胞軟骨である.Köllikerの先例に従って多くの学者は脊索の組織もまたこれに数えているが,一つにはその発生学的な由来が異なること,今ひとつの理由はその化学的組成からして,この説の妥当性は疑われる.発生学的および化学的の両見地からすれば,脊索組織はむしろ上皮性の組織の特殊な一群とすべきものであろう(34頁を比較参照せよ).
Hintzsche, Umbildungen・・・Hyalinknorpel/ Z. mikr.-anat. Forsch., 25. Bd.,1931.
5.骨組織は石灰化した細胞間物質(あるいは原線維間物質)と石灰化しない膠原原線維と,豊富な枝分かれをしてたがいに吻合している骨細胞とからなる.
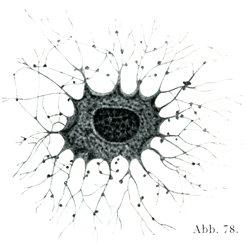
骨細胞は基質の内部の空所内にあるが,この空所の形は骨細胞とその突起の形によく似ている.晒した骨では細胞がなくなってこの空所のみがよく見えるわけで,これを骨小体Knochenkörperchenあるいは骨小腔Knochenlücken oder Knochenhöhlenという(図79).
骨小腔の多くはレンズ形をしていて,長さ13~31µ,幅6~15µ,厚さ4~9µであり,数多くの突起(骨細管Knochenkanälchen)をあらゆる方向に出して,この突起が隣のものとつづく.また骨という器官についてみると,骨細管の一部は骨内の大小いろいろの髄腔や血管の通る空所に通ずるし,また骨の外面にも開いているのである.
骨細胞(図77, 78)は細胞膜をもたないで,その原形質は細かい顆粒を示し,核は楕円に近い形である.細胞体が数多くの突起を出して,これが近在の骨細胞の出す突起と結合している.
v. Ebnerによると骨の原線維間物質は石灰化しており,その中にある骨の原線維じしんは石灰化していないという.他の学者によると骨の原線維も石灰化しているところである.原線維は束をなして集まり,その集束が異なる方向をとってたがいに交叉している.これによって生ずる形象がSharpey-Ebnerの層板現象Lamellenphänomenとう名称で知られている(図161).
これらのはなはだ細かい原線維の他に,それより太くて,骨の表面から骨質の内部にはいっていく線維性結合組織の束がみられるのであって,これはSharpey線維,あるいは穿通線維durchbohrende Fasernとよばれる(図165).
酸を用いて骨組織から無機成分(その集まりを骨土Knochenerdeという)を除去することができる.そのさい残っている有機成分を骨軟骨KnochenknorpelあるいはオッセインOsseinという.これは骨組織の構造をそのままに示しており,軟らかくて曲げることができ,メスで容易に切れる.
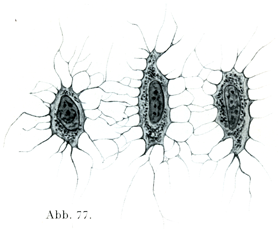
[図77]骨細胞3個(縦に置いて内部までみる.)ネズミRatteの肩甲骨より.×1200. (Ruppericht)
[図78]骨細胞1個(横に置いて内部までみる.)ネズミRatteの肩甲骨より.×2500. (Ruppericht)
骨の無機成分を取り除いてオッセインを得ることができるように,またオッセインを除去して無機成分だけを残すことができる.それは用心深く焼くのであるつまり灰化Verashungであって,そうすると有機成分がこわれてしまう(灰化骨calcinierter Knochen).この灰化骨が骨組織の細かい構造を保っていることはオッセインの場合と同様である.無機質と有機質とが如何なるぐあいに合しいるのか,すなわち単なる混合であるか,もしくは化学的結合の一種なのかがはっきりわかっていない.有機物をすっかり除かれた灰化骨はその形が良く保たれていても,脆くて容易に砕ける.無機成分がオッセインに沈着して初めて,骨組織の丈夫さが得られるのである.
器官としての骨の構造および骨組織の発生については骨学のところで述べる.ここでは人体の発生において骨組織ほおそい時期に初めて現われることだけを指摘しておこう.胎生の第7週に鎖骨がその皮切りをするのである.
6. ゾウゲ質組織Substantia eburnea (Dentin), Zahnbeigewebeは原線維間物質と膠原原線維と特殊な細胞であるゾウゲ細胞Odontoblastenとより成る.
ゾウゲ芽細胞の体はゾウゲ質組織の外にある.細胞体は円柱状あるいは西洋梨状であって,2つの突起をもっている.長い方の突起は歯線維Zahnfaserとしてゾウゲ質組織の中にはいる.そして途中で叉状に2分し,また数多くの側枝を出して,近くにある歯線維の側枝とつづいている.ゾウゲ芽細胞のもついま1つの突起はその下にある歯髄内の結合組織のほうにゆく.核は細胞の底部にあって,楕円に近い形をなし,クロマチンに富み,通常2個の核小体をもっている(von Korff).
歯線維およびその枝はいずれもゾウゲ質を貫くゾウゲ細管Zahnkanalchen oder Dentinröhrcheという細かい管のなかにある.この細管の壁は他の基質に比べて,いっそうつよく光を屈折し,またいっそう固い.そこを歯線維鞘Zahnfaserscheide (Neumann)という.
石灰化した原線維間物質は多量の細かい膠原原線維を有していて,この原線維は歯の表面に平行な方向に走っている.薄い酸で処理するとオッセインとほぼ同じ物質である歯軟骨Zahnknorpelが残る.
[図79]骨小腔(a, a)それから多数の突起が出て,横断されたハヴァース管(b)に通ずることを示す.(Freyによる)

[図80]ゾウゲ細管 ヒトの大臼歯の根を横断して研磨してつくった標本の一部.×350.
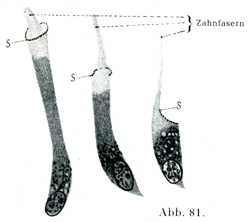
[図81]ゾウゲ芽細胞 ウシの胎児よりとり出したもの.Sは顆粒性の縁で歯線維鞘のはじまりである.(v. Korffによる)
なおゾウゲ質の内部には球間区Interglobularräumeといって,球状の面で境された特異な空所がみられる.これは原線維間物質が石灰化しなかった場所であって,歯線維はそこを貫いて通っている.
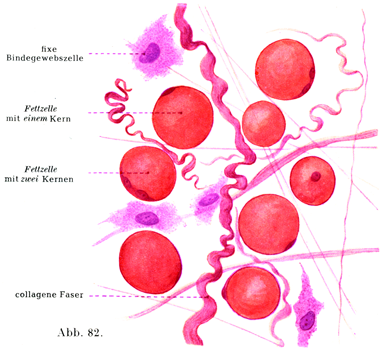
[図82]疎性結合組織内の脂肪細胞.スダンIIIをもって脂肪はオレンジ色に染められている.×300.
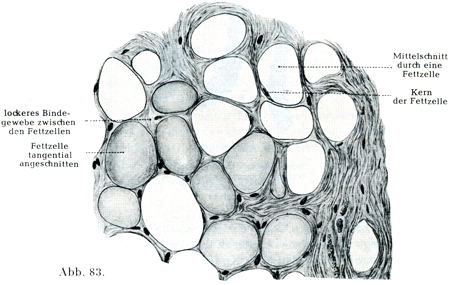
[図83]ヒトの脂肪組織.切片標本.
7. 脂肪組織は脂肪細胞,膠原線維,弾性線維,格子線維,および細胞間(原線維間)物質から成っている.
脂肪細胞は円くて大きい脂肪であり,トリプシンで消化されない丈夫な膜で囲まれている.健康な人だとこの脂肪は脂肪でいっぱいに充たされているので,脂肪球の表面と細胞膜の間に原形質のごく薄い層があるのみである.しかし細胞核のまわりでは原形質がいくらか厚い層をしている.核は楕円に近い形をしており,膜と脂肪球とのあいだに押されて存在するためにやや扁平である(図82,83).2核をもつ脂肪細胞がみられることがあり,時には3核のものある.2つ以上の核があるときは,それらの核がたがいにすぐそばに接近していることと,いろいろ異なる距離で離れていることがある.
新鮮標本を染めないままでみると脂肪細胞の核と原形質はこれらが光学的断層optischer Durchschnittにあるときに限って,はっきりと観察される.その他の一では脂肪滴の輝きがつよくて核の像は全く消されてしまう.脂肪がまだ細胞を完全に充たしていないとき,あるいは消費されて脂肪が減ったときは,その細胞像がいっそうよく理解できる.そういう細胞では核と原形質がはなはだたやすく認められるのである.脂肪の含有が少ないときは脂肪球が1個でなくて,それが2個以上に分かれていることがある(図84,85).
核の内部に1個の脂肪小滴があるときLochkern(有孔核)の像が生ずる.
個々の脂肪細胞のあいだは少量の結合線維があって連ねられている.この結合裾域の誓詞は最初に述べた疎性結合組織と同じものである.
Plenk (Verh. anat. Ges.,1927)によると脂肪細胞は真の細胞膜をもっていないので,細胞膜とみえるものは実はその細胞から分泌された基質とみなすべきものである.但しこの基質は近くにある細胞のまわりの膜とつづいていない.この基質性の被膜のなかに銀好性線維がある.しかし機能的にはこの膜が細胞膜の意味をもっているという.以上の説に反してWassermann (Z. Zellforsch., 26. Bd.,1937)は脂肪細胞の膜は真の細胞膜であると唱えた.その膜のうえに外形質Exoplasmaに由来する細胞の分泌があり,その分泌の中に原線維が含まれているという.
脂肪細胞のでき方は:1. 普通の結合組織細胞が脂肪を摂取し,あるいは製造することによって,結合組織の内部でかなり散在した状態であらわれる.2. 脂肪器官Fettorganeともいうべき特別な器官をなしてできてくる.これはKöllikerがPrimitivorgane der Fettkeimleger(脂肪のもとをなす倉庫)とよんだものである.これは脂肪をもっていることも,もっていないこともあるが,何しろ特殊な脂肪組織である.これらの脂肪器官の出発点はやはり若い結合組織細胞であるが,その集まりが灰赤色の小葉をなしてあらわれる.それをなす細胞は丸みをおびた多角形で,細胞膜を欠き,顆粒をもたないで,きれいな核をもっている.形質細胞と似ているのである.これらの事実はWassermannの研究(Z. Zellforsch., 3. Bd.,1926)によっても確かめられたが,Wassermannはこうゆう脂肪器官を結合組織とは別の新しい形成物と考えたのである(z. Kreislaufforsch., 23. Jahrg.,1931).
身体における脂肪組織の意味ははなはだ大きい.栄養の目的の他に外力に対して体を保護する蒲団であり,熱を保つための被いもものであり,鋭い高まりやへこみをなくしたり,隙間をみたす詰めものとして大きい役目をなしている.詰めものの例としては黄色骨髄がある.栄養のより人体では脂肪組織の目方は15キログラムか,それ以上にまで達する.
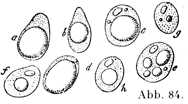
[図84]脂肪組織の退行Rückbildung (Freyによる).やせた衰えたヒトの死体の皮下結合組織より得たもの.aは大きい脂肪滴をもつもの,bは小さい脂肪滴をもつもの,cとdは核がみえるものも,eはばらばらになった脂肪滴をもつ脂肪,fは小さい脂肪滴を1つだけもつもの,gはほとんど脂肪を有しないもの,hは脂肪がなくて,蛋白質様の物質の1滴を細胞体の部分に有するものである.
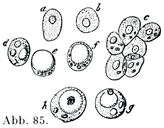
[図85]脂肪組織の発生Entwicklung (Freyにる.)10インチの長さのヒツジ胎児の腎臓のまわりにある脂肪組織の細胞.aとbはまだ始部を有たない細胞,cは脂肪を有たない細胞の集まり,d~hは細胞体の一部を占めて細胞がいろいろの量にたまってゆく各段階を示してある.
8. リンパ様組織lymphoides (cytogenes, adenoid. es, retikuläres)Gewebeはリンパ細網組織lymphoretikuläres Gewebeという名前がいっそう適切であるが,大部分が円い小さい細胞(リンパ球)より成り,結合組織性の線維が網状をなして支え,また少量の細胞間物質がある.
リンパ球は小さい球形の脂肪である.この細胞の大部分を占めて球形の核があり,声は密なクロマチン網を示す.原形質は核のまわりにせまい縁をなすのみである.この小さい細胞の他に数はいっそう少ないがもっと大きい核をもち,細胞体もいっそうよく発達している細胞がみられる.これらの細胞は細胞線維Retikulumfasernという結合組織線維がつくる支柱の網の目に存在する.この網はリンパ球があまり多数であるため,それに被いかくされている.切片標本をつくって,細胞成分を筆で掃いのけたり,切片を揺り動かして細胞を除くか,あるいは特殊染色で細網線維をそめることによって,初めて支柱をなす網の像をはっきりみることができる.細網組織retikuläres Gewebeという名称はこの像に基づくのである.
リンパ様組織は体内にはなはだ多くの存在する.特にこれはリンパおよび血液をつくる多くの重要な器管にある.すなわち数多いリンパ節,孤立および集合リンパ小節,所々にある扁桃,胸腺,脾臓の中にある.なお粘膜の定まった場所,舌や腸や呼吸器系の粘膜および結膜にもみられる.
これらすべての場所でリンパ様組織はリンパ球や白血球の発生するところBrutstätteとみなさえるのである.扁桃の組織や孤立および集合リンパ節の組織は比較的近年の研究によれば上皮細胞とリンパ球とが密接に混じり合ったものから生じたといわれる.そのためにリンパ上皮組織lumphoepitheliales Gewebeとよばれるのである.
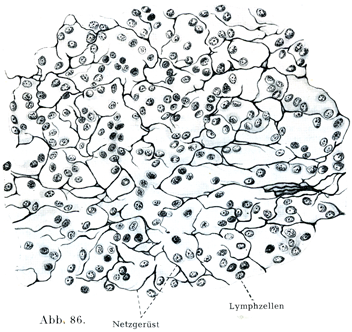
[図86]リンパ球をもつ細網組織.ヒトのリンパ節より.×600.
9. 色素結合組織pigmentiertes Bindegewebeは色素結合組織細胞すなわち枝分かれをした細胞体で,これが色素顆粒でみたされている結合組織細胞を多数にもったことが特徴であって,そのほかに膠原線維と弾性線維また色素顆粒をもたない別の種類の結合組織細胞ならびに少量の細胞間(原線維間)物質がある.
この細胞は結合組織細胞1)の細胞体に褐色の色素顆粒がつくられるか,あるいは取り込まれることによって生ずる.細胞体が色素顆粒で全く充たされていることがある.しかし核はそのときも色素顆粒を有しないで,それを被いかくそうとする色素の中から明るい小胞としてみえるのである.はなはだ普通に色素細胞は枝分かれして,星形を呈している.
色素結合組織細胞の存在は人間では眼(眼球中膜と強膜),クモ膜,および皮膚に限られている.
1)色素上皮細胞pigmentierte Epithelzellen, 色素結合組織細胞pigmentierte Bindegewebzellen, 色素神経細胞pigmentiete Nervenzellen, 色素筋細胞と色素筋線維 pigmentierte Muskelzellen und-fasernがある.色素は常に組織細胞Gewebezellenの内部にある.色素上皮細胞は有色人種の皮膚に広い範囲にわたって存在する.白色人種では陰茎,陰嚢,肛門付近,腋窩,眼瞼の皮膚や眼の結膜にそれがある.色素は主として表皮の下部の細胞層にある.色素結合組織細胞は体じゅうどこにも存在し得るが,最もその数の多いのが眼球中膜Tunica media oculi(すなわち脈絡膜と毛様体と虹彩)であり,それについで多いのはすぐ上に述べた皮膚の一部で色素に富む場所である.動物では色素顆粒をもつ軟骨細胞pigmentierte Knorpelzellenも存在する.まだ瞳孔散大筋M. dilatator pupillaeは色素筋細胞よりできている.色素神経細胞は中枢神経系の特別の場所にある(黒核Nucleus niger,青斑Locus caeruleus).比較的年配の人の神経細胞には正常の現象と色素が存在する.(原著註)
10. 内皮組織は脈管の内面を被うごく薄く平たい細胞から成っている.個々の細胞は扁平で,その輪郭は多くの場合不規則な波状である図88,89).ただ2,3の個所,たとえば前眼房の内皮では直線的の境をもった規則正しい五角形あるいは六角形の板状の細胞がならんでいる.モザイック状に配列した細胞の縁のあいだには細胞間橋と細胞間隙とがあり,後者を通って液が通じ,遊走細胞が移動している.内皮細胞の表面は,つまり内皮が囲むところの空所に向かっている表面であるが,個々は平滑であって,比較的固い1層をなしている.扁平な楕円形の細胞核があって,局部的にそこだけ細胞体が高くなっていることが少なくない.
脾臓の静脈性毛細管(いわゆる脾洞Milzsinusのことである.(小川鼎三))の内皮細胞(II巻をみよ)は特別な形をしている.それは細長くて,両端が尖ってい終わる棒の形であって(杆状細胞Stabzellen),その核は管の内腔の方につよく突出している.
肝臓の小葉内の毛細管の内皮細胞もまた特異性を示す.その細胞境界が銀液で処理しても検出することができないのであって,従っておそらく合胞体Syncytiumなのである.
11. 胎児性結合組織embryonales Bindegewebeは初めはただ,たくさん枝分かれをした星形の細胞がその枝でたがいに結合して(すなわち吻合して)おり,その細胞のあいだにムチン(粘液素)をもつ膠様の物質があるのみである(図63).その細胞の核は楕円に近い形か,あるいは細胞体の形に応じて不規則な形をしている.
胎児の結合組織がこの形態にとどまるのは短い期間のみであって,まもなく細胞の中にも外にも膠原性および弾性の原線維が各々の場所によって様子はちがうがあらわれる.しかし細胞間の膠様物質は何しろ後ろまで豊富に存在しているので,出産の時でも臍帯の組織はそのときはなはだしく線維に富んでいるにかかわらず,なおジェリー様の性質を示すのである(ワルトン軟肉Whartonsche Sulze).また胎児の結合組織が永くみられる場所としては内耳の膜迷路のまわりである.これが液化していわゆる外リンパ腔perilymphatische Räaumeが生ずる.
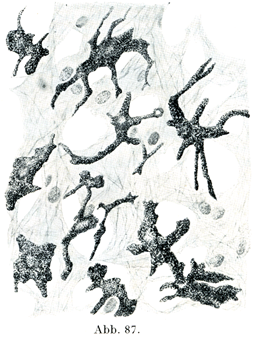
[図87]色素結合組織細胞 眼の脈絡外層Stratum perichoriodeum. ×600.
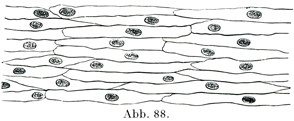
[図88]脳軟膜の動脈の内皮.硝酸銀で処理して細胞の境をあらわしてある.×300.
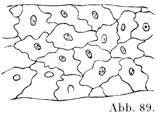
[図89]リンパ管の内皮.モルモットの腸の筋層より.×240. (Auerbachによる.)
細網内皮系
Aschoffは結合および支持組織に属するいろいろな種類の細胞を一とまとめにして細網内皮系retikuloendotheliales System (RES. と略記することがある)とよんだ.これらの細胞に共通する性質は陰電気をおびる膠質や,体内および体外に由来する有形要素に対して特別につよい貯蔵能力を持つことである.この貯蔵はまず顆粒の形でおこなわれて,ついでその顆粒がさらに変化せられるのである.
結合および支持組織の細胞は細網内皮系に属しないものでも貯蔵能力があるが,しかしそれは色素を多量にははたらかせたときに限り,またごく細かい顆粒の形でのみおこるのである.じっさい,動物体の細胞はどれでも或る条件のもとでは異物をその細胞体の中に取り込むことができるであろう.そこで,細網内皮系に属するか否かの区別となる標準は食作用Phagocytoseの強さと頻度なのである.
狭義の細網内皮系は脾臓やリンパ節やそのほかのリンパ様組織の細網細胞,ならびに脾臓の毛細管性静脈(すなわち脾洞),リンパ節のリンパ洞,肝小葉や骨髄や腎上体皮質や下垂体の内皮細胞からなるのである.
広義の細網内皮系に属するものとしては以上のほかに,結合組織で運動している細胞,脾髄の細胞,循環している血液のなかの単核細胞Monocyten, 毛細血管の壁のまわりにある外膜細胞Pericyten oder Adventitiazellenである.
細網内皮系に属しないものは普通の血管やリンパ管の内皮細胞,固定結合組織細胞(すなわち線維細胞),形質細胞,肥満細胞,胸腺の(上皮性起原をもつ)細網細胞,神経系のグリア細胞,硬膜下腔や軟膜腔の内皮細胞,胸膜腔と腹膜腔と胸膜腔の内面を被っている上皮細胞である.
血液とリンパBlut und Lymphe
血液は細胞のみからなり,その間に豊富な細胞間物質をもつ,結合および支持組織の一種とみなすことができる.それゆえ血液には2つの主な成分区別さえる.1. 液性成分すなわち血漿Plasma s. Liquor sanguinisと2. 有形成分すなわち血球corposcula sanguinis, Blutkörperchenである.血漿は凝固という現象によって2つの成分に分かれる.a)固型で線維顆粒性fadigkorningである線維素Fibrinとb)液性で黄色みを帯びて透明な血清Serumである.有形成分にはa)赤血球Erythrocyten, rote Blutkörperchen,b)無色血球farblose Blutkörperchen,これは不適切ながら白血球Leukocytenともよばれる.c)血小板Thrombocyten, Blutplattechen,d)血塵Hamokonien, Blutstäubchen,これはおそらく特別のものではなく別の由来をもつ蛋白質であろう.また血塵の一部は脂肪小滴である.
次の表がその全体をも通すのに便利であろう.
|
血液 |
血漿 |
線維素 |
|
血清 |
||
|
血球 |
赤血球 |
|
|
白血球 |
||
|
血小板[血塵] |
血液は全体としてみると1つの器官である.生きているあいだはたえず管の形の通路のなかを経めぐってい運動している液状の器官である.そえでも1つの器官であるに相違ない.しかしここでは器官としての血液を論ずる必要がないので,組織としての血液についてだけ述べる.
成人における血液の全量はBischoffによると体重の1/13,その後の研究によって体重の1/15~1/20とされている.血液の比重は1.050~1.060(弾性では1.055~1.060,女では1.050~1.056),塩からい味と特有なにおいをもち,暗赤あるいは明るい赤色を呈している.その78~79%は水で,21=22%が乾固物質である.また有形成分が全量の45~50%を占める.Hoppe-Seylerによると,人間の血液の血漿量は67.90%であって,そのうち7.07%が乾固物質,60.83%が水であるという.
1. 血漿Liquor sanguinisは形をもっていない.しかし一定の事情のもとで,それは特に血液を血管の外にとり出したときであるが,凝固Gerinnungという名前で呼ばれる.興味ふかい変化が血漿におこる.それまで透明無色であった液のなかに細く明るい糸状のもの(線維素糸Fibrinfäden)があらわれて,これが急速増加して,ほかの有形成分を閉じこめてしまう.このようにしてはなはだきれいな網ができるのである.場合によってはこの糸のできかたが不規則であって,しまいには密にもつれてフェルトのようなものが生ずる.その物質が血液の線維索Fibrinとよばれるのである.(凝固現象の詳細については54頁を参照のこと).そのときに残っている液体(有形成分を全くもっていない)が血清Serum sanguinisである.血液の有形成分と線維素をもつ部分いわゆる血餅Placenta sanguinis, Blukuchenである.血液が凝固する前に棒をもってこれをかきまわすと,線維索は白い線維状の塊として,ごくわずかの血球のみを伴って,分離される.そのときに残っている部分は脱線維血液defibriniertes Blutとよばれる.
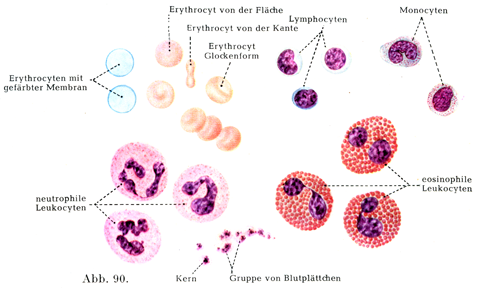
[図90]ヒトの血液 ×1000. 乾燥標本を染めて赤血球,白血球,血小板の形状を外観を示す.単核球の図はSchulten, H., Klinische Hämatologie. V. Aufl.1953. Thieme, Stuttgartより借用した.
2. 赤血球Eryghrocytenは出生後の生活では核をもたない,両凹の円板状で,その縁は丸みをおびている.円板の陥凹はへこみDelleとよばれる.赤血球の色は顕微鏡の舌では黄色をおびている.肉眼で見ると赤い.その色は特別な蛋白体であるヘモグロビンHämoglobinに基くのである.円板の直径は平均7.5µで,少数のものは8~12µに達して,巨大赤血球Megalocytenをなす.ほかのものは矮小であって2~4µに過ぎず,これを小赤血球Mikrocytenという.厚さは縁のところで2.5µ,中央部で1.8~2µ(Hayem)である.赤血球は軟らかくて曲がりやすい.しかし弾性にとむ.また標本の中では赤血球が列をなしてならぶ傾向があってその広い方の面をもって相接している.これによっていわゆる銭包み配列Geldrollen-Anordnung(図91)が生ずる.しかしこれは血液層がある一定の厚さをもっているときにのみ起こる現象である.
赤血球は液性の内容物すなわちヘモグロビンをかこむ.細胞膜をもっている.また赤血球ははなはだ敏感なものであって,その形や性質が種々雑多な影響のもとで変化する.
水分が蒸発したり,あるいは塩類や糖類を加えることによって,浸透圧が高まると赤血球は縮む.そのさい尖った小突起ができて,赤血球は桑の実Mauleerあるいは朝鮮アサガオの実Stechapfelに似た形となる.浸透圧の減少は膨化,膜の破れ,ヘモグロビンの流出を引きおこす.そのとき細胞膜は容易に認めがたい陰のうすいもの(血影Blutschattenという)として残っているが,ヨード液を加えるとそれがはっきりとみえるのである.
細胞膜は透明無色であって,生の標本ではみえないで,粘着力がある.メチールヴィオレットによって新鮮標本でも(Schäfer), また固定保存された標本でも(Deetjen), 細胞膜を染めてあらわすことができる.この膜はリポイドの外層と蛋白質の内層とから成っている.赤血球の内部に特別な細胞形質の構造は存在しないようである.むしろヘモグロビンという液をもって充たされた中空の球であって,その壁がいま述べた細胞膜であると考えて良いであろう.
赤血球の少数(3.15%)のものは適宜な染色をほどこした場合,その内部に細かい顆粒や糸の構造を示すのであって,これは網状赤血球Retikulocytenとよばれる.おそらくこれは若い赤血球であろう.
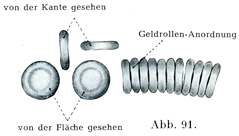
[図91]ヒトの赤血球.×1200

[図92]ヒトの赤血球.朝鮮アサガオの実の形(金米糠状).
比較解剖学:哺乳動物の赤血球は人のものと非常によく似ている.最も大きさはいろいろと異なる.象やセイウチや貧歯類の赤血球は人のより大きい.犬のそれは7.3µの直径である.諸種の動物の赤血球の大きさについての研究結果を記した大きい表ができている.ラクダ類の諸動物の赤血球はやはり核を有しないが,形が楕円である.ほ乳里以外の脊椎動物では赤血球は楕円形で,核をもっている.ヤツメウナギのそれは核をもっていて円い輪郭の円板である.これらの関係について図93が示している.
動物界には2種の違った血液がある.鉄の血Eisenblutの銅の血Kupferblutとである.前者はヘモグロビンをもち,空気に触れたとき赤くなり、後者はヘモチアニンHämocyaninをふくんでいて、空気に触れたとき青くなる.これは軟体動物や節足動物にある.鉄の地は脊椎動物以外でも,かなり多くの蠕形運動や軟体動物にみられるのである.
人および哺乳動物でも胎児の或る時期までは赤血球のすべてが核をもっていたのであって,そのときは有核赤血球の有糸分裂が数多くみられ,この分裂は赤血球が循環しながらおこなわれる.発生がさらに進むと有核赤血球の量が減る.人の血液はすでに胎生第4月には無核赤血球がはなはだ多くなり,第6月にはもはや有核赤血球はごく少数となり,第7月より後は核の内戚血球のみが存在する.その後の時期でも一定の病気の時には有核赤血球があらわれる.
赤血球の寿命はいろいろと異なるのが短いので14日,永いので200日といわれる.平均の寿命はRuhenstrothとBauerによれば45~60日に過ぎないという.おそらく順庵している血液の中で死滅するのがあるだろうが,そのほかに,肝臓や脾臓の中,またたぶん骨髄の中でも死滅する.骨髄では一定の細胞が赤血球摂取して,これを変化させるのである.
3. 白血球Leukocyten, farblose Blutkörperche(図90)は細胞膜をもたない細胞であって,顆粒性の原形質と1個あるいはそれ以上の数の核より成っている.
休止の状態では球形であり,4~14µの直径をもっている.アメーバ様の運動をするので,従っていろいろと変わった形をとる(図94).
細胞の大きさ,核の形,ならびに細胞体の構造やその染色性によって白血球に5種が区別されている(図90).その第1と第2の種類は単核白血球uninucleäre Leukocytenとして,その他のものすなわち多核白血球multinucleare Leukocytenと比較されるのである.白血球のあらゆる種類はおそらく1つの共通の原形かあできてくるのであって,その原形としてはリンパ球が考えられる.
a)リンパ球Lymphocytenはたいてい赤血球くらいの大きさであって,つよく染まる比較的に大きい球形の核と,この核をとりまく原形質のせまい薄い層をもっている.
b)単核球MonocytenはEhrlichにより移行型Übergangsformenとよばれたもので,正常の血液細胞としては最も大きいものであり,20µに達するのがある.単核球はリンパ球に似ているが,大きさがいろいろであって,染色性のよわい1個の大きな核を有し,これが時としては分葉形である.
c)中好性白血球neutrophile Leukocytenは赤血球のほとんど2倍ほどの大きさである.核はいわゆる多形polymorphすなわちいろいろと異なる形である.馬蹄状,クローバの葉の形,あるいは不規則な形をして,いくつかの部分に分かれており,つよく染色する密な核材をもっている.細胞体は密集した細かい顆粒をもっていて,この顆粒は酸性の色素にも塩基性の色素にもよく染まらないで,それらを混ぜてつくった色素液の中で,一方に偏しない色調に染まるのである.
d)酸好性白血球acidophile (eosino-oder oxyphile) Leukocytenは中好性白血球と同じぐらいの大きさで,やはり多形の核をもっている.この核もいくつかの辺に分かれてるのがしばしばである.細胞体はつよく輝くところの粗大な顆粒で密にみたされていて,この顆粒がエオシンやフクシンのような酸性色素をむさぼるごとく摂取するのである.
e)塩基好性白血球basophile Leukocytenは数がはなはだ少なくて,細胞体は大きい顆粒に富むが,この顆粒が比較的に少量であることもある.顆粒は塩基性アニリン色素に染まり,その大きさと形がはなはだ不規則である.この細胞の核は多形である.
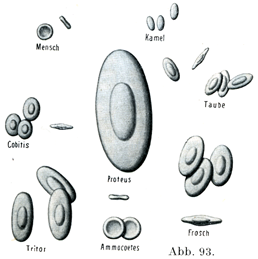
[図93]ヒトおよび諸種の動物の赤血球を同じ拡大で示す.(Freyによる.) Mensch・・・ヒト,Kamel・・・ラクダ,Cobitis・・・シマドジョウ(硬骨魚類),Taube・・・ハト,Triton・・・イモリ(有尾両棲類),Proteus・・・ヨーロッパの洞窩に住む盲目の有尾両棲類,Frosch・・・カエル,Ammocoetes・・・スナヤツメ(円口類)の子供.
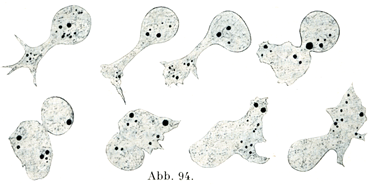
[図94]アメーバ様の運動 カエルの白血球の1個が10分間に示した形状の変化.24時間前にそのカエルの背部リンパ嚢に墨汁を注射したので,墨の粒が白血球の中に取り込まれて大小の黒い点としてみえる.
上に述べた3種の顆粒をもつ白血球(c,d,e)を総括して顆粒白血球Granulocytenという.核の形によって分けると棒状核stabkeringと分節核segmentkernigの顆粒白血球とがある.核がいくつかの片に分かれているとはおそらくその白血球が古くなったことを示すのである.
白血球の有糸分裂はSpronckによると哺乳動物の循環している血液の中でも見られるのであって,その頻度は1000の白血球におよそ2つである.白血球の数が増加するための,絶え間ないいま一つの源をなしているのがリンパであって,これは本書ですでにリンパ球の生成するところとして述べた諸器官から新しい白血球を血液の中に絶えずみちびき入れているのである.
上述のことからわかるように,血液およびリンパのもつ白血球,リンパ球やリンパ性の細胞は同じ由来のものである.骨髄もまたほかの流水のもののほかにこの類の細胞をもっている.すなわち骨髄細胞Markzellenがそれである.その他の組織ではこれと同類のものが遊走細胞Wanderzellenとよばえて散在している.遊走細胞の少なくとも一部は血管から外に出てきたか,あるいは白血球じしんがその成生した所から直接にやってきたものである.遊走細胞のほかの一部,すなわち単核球はおそらく固定結合組織細胞に由来する.
遊走細胞は上皮の細胞間にはいってゆくこともできるのであって,かくして上皮の表面に達し,あるいは上皮層の内部で壊れてしまう.消化管の全体で遊走細胞が上皮の表面に達する(Stöhr).それゆえ唾液の中にも入っており,ここでは唾液小体Speichelkörpercheとよばれて,唾液のはたらきの一翼を担っている.遊走細胞および一般的に云って白血球は,それらが如何なる所にあっても,必要な場合には食細胞Freßzellen, Phagocyten (Metschnikoff)としてはたらきうるのである.すなわち小さい警備器官として作用して,使い古したものを身体から取り除くことを手伝ったり,病気を起こす微生物を摂取して消化したり,細胞の壊れてできた屑を身近らの細胞体内に取り入れて,それを掃除したりする.また白血球が血管からどっと外にでてゆき,それも内皮細胞の間を通って外に出たものが膿Eiterの主成分をなすのである(膿小体Eiterkörperchen).両棲類の遊走細胞では中心小体がいつまでも消えないで認められたのである(図10).ヒトの白血球では2個の中心子がたがいに密接してならんでいる.
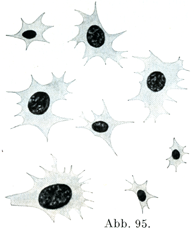
[図95]血小板 ヒトの血液,Deetjenの考案した液を用いた.1%のオスミウム酸で固定,ヘマトキシリンで染色.×2000.

[図96]血小板 ヒトの新鮮な血液より直ちに固定した.面の方向および角の方向よりみたもの.×2000.
4. 血小板Blutplättchen, Torombocyten (Dekhuyzen)は小さいもので,1~3µの大きさであり,一部の学者(Deetjen, Kopsch等)によれば核をもつ細胞である.循環している血液の中では血相板の形はその面の方向からみれば円く,角の方向からみると砥石状である(図96).新鮮血液を固定した標本でみると若干の血小板は1つあるいは2つの細い突起を出している.血小板の比重は血漿のそれより低いので,生の血液標本ではそれが上方の層に集まるために,顕微鏡の焦点を被いガラスの下面に合わせると,それが明るく輝く点状のものとして容易に認められる.血小板は非常に不安定なものであって,そのまわりの血漿がごくわずかの変化を示しただけで,これが害されると.そのとき血小板はたがいに粘着し,また近くにある対象物に固く付着して,核が溶けて原形質が泡のようなものにかわって破壊するのである.この現象が血漿の凝固を引き起こし,またそれを条件づけるものである.
Deetjenの方法によってつくられた新鮮標本では血小板の各区が緑色をおびた輝く点状のものとして認めあれる.その原形質が尖った突起をのばし,これがふたたびひっこめられる.
ごくわずかであるが,位置の移動されも血小板で観察されている.図95は血小板がどういうぐあいにみえるか,その大きさがどれくらいちがうかを示すのである.
人の血液の血小板が下等動物のやはり血小板と呼ばれる有核で同じ機能を持つ細胞と相同のものであるかどうかは未だ決定していないが,相同であることの可能性が大きい.
Deetjenやその他の人々によって核と考えられた細胞内容物は顆粒部Granulomerとよばえていて,多くの学者はこれを核とみなしていない.しかし組織化学的に核反応が陽性である.J. H. Wright (Publ. Mass. Gen. Hospital. Boston, Vol. III., 1910)は血小板が造血器官(骨髄,脾臓)内の巨大細胞の細胞体からちぎれて生ずることを発表した.そのさい顆粒部は細胞体のもつ顆粒に由来するのであって,核から生ずるのではないと云った.Pfuhl (Z. Anat. Entw.110. Bd.,1940)によると血小板はトリパンブラウによって“すっかり”durchweg染まるので,生きている細胞ではない,生きている細胞だとそんな染まり方をしないというのである.
5. 血塵Blutstaubchen (Hämokonien, Muller)および脂肪小滴Fetttröpfchen. .血漿内にいろいろと異なる量に存在する顆粒および顆粒集団はMüllerが考えたのと違って,独立の成分とはおもえないものである.生気のとぼしい外観を呈していて,最も大きい構造物が壊れてできた破片であろうかとの考えをおこさせるのである.
血漿の中にある脂肪小滴は乳ビやリンパの膨らむ美貌が血液の中に注がれたものとるすのが,最も考えやすいことである.
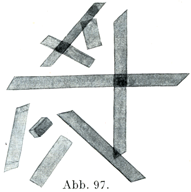
[図97]ヒトのヘモグロビン結晶(Schäferの図を利用した.)
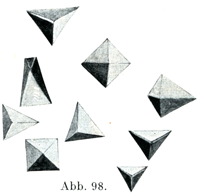
[図98]モルモットのヘモグロビン結晶(Schäferの図を利用した.)

[図99]ヒトのヘモグロビン結晶(Schäferの図を利用した.)
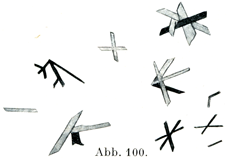
[図100]クロールヘミン結晶(Teichmann結晶)(Schäferの図を利用した.)
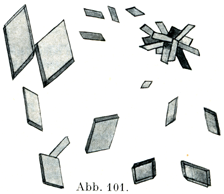
[図101]ヒトのヘマトイディン結晶(Schäferの図を利用した.)
6. 血結晶Blutkristalle:次のものの結晶が知られている.1. ヘモグロビンHämoglobin, 2. メトヘモグロビンMethämoglobin, 3. パラヘモグロビンParahämoglobin,4. オキシヘミンOxyhaminとクロールヘミンChlorhämin, 5. ヘモクロモーゲンHämatoporphyrin, 6. ヘマトボルフィリンHämatoidin, 7. ヘマトイディンHämatoidin,8. シャルコー・ライデン結晶Charcot-Leydensche Kristalle,9. 血清結晶Blutserumkristalle,10. フォルマリン色素結晶Formalinpigmentkristalle.
そのうちで最もよく知られているのはヘモグロビン結晶,クロールヘミン結晶,ヘマトイディン結晶である.
ヘモグロビンは動物の各種にそれぞれ特有な形の結晶をなす.人では柱状(図97),モルモットでは四面体(図98),リスでは六角形の板(図99)である.
クロールヘミン(前には単にヘミンHäminといった)はその化学的に純粋なものがタイヒマンTeichmann結晶のなかにある.この結晶はよくで来たときは菱形の小板や柱をなして,複屈折性である.まずくできたときは卵形,大支の粒の形や文章の節の符号の形(§)をしている.
とくに注意を要することはタイヒマン結晶の形がヘモグロビン結晶と大いに違って,はなはだ多くの種類の動物を通じて一致していることであって,無脊髄動物のクロールヘミンでも脊椎動物のそれと結晶の形では決して区別できないのである.
オキシヘミン(前にはヘマチンHämatinといった)はクロールヘミンにアルカリをはたらかせるとできる.オキシヘミンとクロールヘミンはポルフィリンと鉄の化合物である.
ヘマトイディン(R. Virchow)は血管の外にできた血液塊が生活体の中で徐々に濃く固くなったとき,どこででもできる(例えば脳の内部,卵巣の黄体の中,皮下結合組織の中).オレンジ色の菱形の小板である(図101).
シャルコー・ライデンCharcot-Leyden結晶は喘息結晶Asthmakristalleともいい,ある種の病気の時に,こわれた白血球がともになって生ずる.長くのびた重複ピラミッド形で,その縁がまっすぐで,その角はとがっている.
メトヘモグロビンは酸素とヘモグロビンが固くくっついたものであって,その結晶は褐赤色の針状,稜柱状などである.パラヘモグロビンはオキシヘモグロビンより生ずる.これは水に不溶性である.ヘモクロモーゲンはヘモグロビンを構成する一部で,有色の原子群であり,鉄を含む.これに反してヘマトポルフィリンはやはり赤いが,鉄を有しない.後者はある種の中毒の時生体内にできる.血清アルブミンの結晶はGürberによって初めて六角形の稜柱の形で得られたものである.フォルマリン色素結晶は血液を含む期間があまり新鮮でない状態でかなり長くフォルマリン液に貯えられたときに,ヘモグロビンから変化して生ずるものである.-Kobert, H. U., Mitteil. aus d. Institut für physiol. Chemie usw. Rostock. Leipzig,1900.
血球の数の問題ははなはだ重要である.それが血液の正常を知るのにしっかっりした基準をあたえるからである.血液の有形成分3種の比は1L. :666 E. : 66 T. である.すなわち1個の白血球に対して赤血球666個と血小板66個がある.血漿と有形成分との関係は次のごとくで,1立方ミリの血液が男では500万個の赤血球,5千ないし1万の白血球,50万の血小板をもっている.
赤血球の数は性,年齢,体質,日時によって変質する.1cmmの血液は女では450万,男では500万の赤血球がある.この差異は思春期以後に初めてあらわれる.
新生児では1cmmの血液に690万の赤血球があることがわかっている.10才までのあいだにその数が450万まで下がり,その次の10年のあいだに500万に増すが,女では思春期までに450万に下るのに反して,男では高年まで500万という数が変わらずにつづくのである.1昼夜のあいだにあらわれる変動は食物を摂る時間に応じている.取り込まれた液体の量によってその数がいろいろなぐあいに変わるのであって,それは当然に血液の含水量が増減することによっておこるわけである.
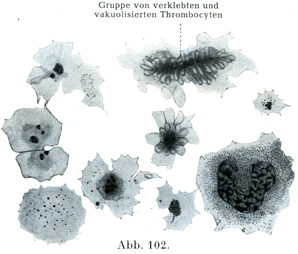
[図102]ヒトの血液の血小板の変化 血液凝固の初期である.右下方に1個の白血球.×1600.
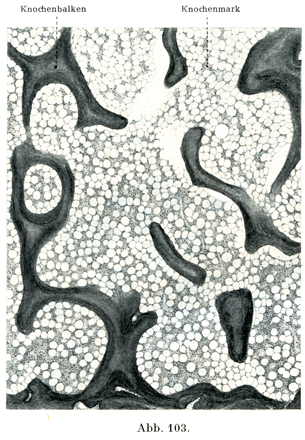
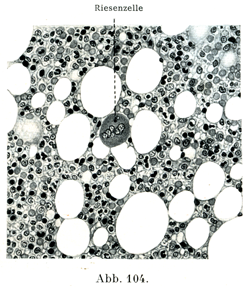
[図103,104]骨髄 ヒトの胸骨. 図103は×30, 図104は×300の拡大. 明るい円形の隙間は脂肪細胞である.
白血球の数は年齢による差異のほかの著しい個体的差異を示し,また1日のうちの時刻によってもある程度変動する.新生児では生後のいく日かは1cmmに18,000の白血球が数えられた.10才までの間にはその数は12,900となる.中年の人では平均して1cmmに7, 680であり,すなわち平均値を7, 500と仮定すると,666個の赤血球に対して白血球が1個の割となる.白血球の各種の割合は:好塩基好性が0.5%,中好性が65~70%,リンパ球が25%,酸好性白血球と単核球を合わせて5~10%である.-Arneth, J., Qualittive Blutlehre und Blutkrankheiten. Leipzig.1942.
血小板の数を決定することは,このものが不安定な構造をもち,また集団をなす傾向があるために,はなはだ困難である.1cmmにおける血小板の数はAffanassiewによると200,000~3000,000,Fusariによると180,000~250,000,Preussによれば500,000,BrodieとRussel, Prattによれば400,000~500,000である.
血液の凝固を顕微鏡でしらべると,初めの4~5分間に血小板が特異な変化を起こし(図102),それは細胞体にも核にもおこるのである(Fr. Kopsch).すなわち細胞体が著しく増大し,核は壊れていくつかの顆粒に分かれる.細胞体の増大は液胞形成によっておこるので,血漿案の集団が泡状の観を呈するのである.
この変化が経過した後に初めて線維索の形成がはじまり,それとともに血漿の凝固,線維索と血清との分離がはじまる.線維索は細かい糸の形で,その一部は平滑であり,一部は顆粒をもっている.この細枝が放線状にすすんであらゆる有形成分に付着している.かくして線維索の糸でできたきれいな,眼の細かい網が生じて,この網が血小板の壊れたものやその他の有形成分をつつんでします.その後ではこの糸の塊が縮んで,そのために血清が押しだされる.凝固に要する時間は温度によって異なり,また1日のうち時刻によってお変動する(Bürker).温度が高いほど凝固に要する時間は短い.しかし別々の人でそれぞれ別の日に測ってみても,温度がほぼ同じで,それに時刻が同じであれば凝固に要する時間がほとんど同じである(Bürker).MorawitzおよびBürkerの研究は凝固が血小板の破壊と深いつながりをもつことを立証した.このことはBizzozeroのその他の人々がすでにその以前から主張してきたのである.
血球の新生Bildung neuer Blutkorperchen
その寿命が終わったために消失したか,もしくはその他の原因で失われた血液の有効成分の補充はいろいろな場所でなされるのである.
白血球はすべてのリンパ性の器官(リンパ節,脾臓,骨髄)において既存のリンパ細胞が分裂して,これがリンパの流れを介して血液中に洗い出されることによって生ずる.
血小板はWright(55頁)によれば骨髄の巨大細胞から,Watzka(1938)によれば脾臓の巨大細胞や同じく細網細胞や脾洞の内皮細胞(杆状細胞Stabzellen)からできるという.
赤血球の形成は赤色骨髄でおこる.それもおそらくは血管の内部においてである.赤血球ができる場所と様式,とくにはじめに存在していた核がどういうぐあいにしてなくなる,またその他の点の詳細についていまでもまだ一致した見解が得られていない.赤色骨髄には多核の巨大細胞,白血球,骨髄細胞が存在する.骨髄細胞にはヘモグロビンを有たないのと,有っているのとがあって,後者の核が放射状の構造をもち,有糸分裂を示し,またその細胞の大きさがはなはだまちまちである(6~9µ).これが白血球の前身であって,赤芽細胞Erythroblastenとよばれる.循環する血液の中には白血球としてはいる前にその核が失われる(図104,105).
一部の学者(Ehrlich, Maximow)によれると核はまるごと細胞体の外にだされる.他の学者(Neumass, Kölliker, Grawitz)によると核は細胞の内部でだんだんと消えてしまう.
ヘモグロビンを有たない骨髄細胞は白血球の前身である.これが幼若な形から分節核をもつ白血球にまで発達する(図105).発達のあらゆる段階を示す,いろいろな形状の細胞がむらがって,細網組織の中に多数の(多少の差はあるが)脂肪細胞とともに存在している.細網組織を構成している細網細胞はその他の骨髄細胞よりも大形である.
白血球は血液内の細胞としてはわずかに1000分の2ほどにすぎないが,造血器官としての骨髄ではおよそ70%が白血球の前身であり,わずか30%が赤血球の前身である.-Schulten, H., Lehrbuch der klinischen Hämatologie. V. Aufl.1953. George Thieme Verlag, Stutgart.
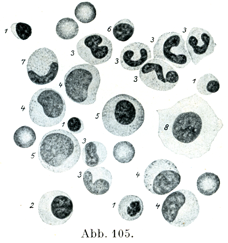
[図105]正常の骨髄にみられる種々の細胞形 (H. Schulten. Leipzig 1937, G. Thiemeによる).1. 古い方の赤芽細胞,2. 若い方の赤芽細胞,3. 中好性白血球,4と5.白血球の前身:そのうち4は骨髄細胞Myelocyten, 5. は前骨髄細胞Promyelocyten, 6. リンパ球,7. 酸好性白血球,8. 細網細胞.
リンパLymphe
リンパは血液から赤血球を取り除いたものによく似ている.それは透明な無色の液,あるいはまた乳様の液としてみえる.その乳のようにな性質はごく細かい脂肪小滴を無数にもつことによるのである.有形成分としてはその他に白血球,リンパ球,リンパ性の細胞があり,一部の学者によると赤血球の前身である赤芽細胞も存在する.リンパ球はすでに記した数多くのリンパ球生成個所から由来するのであって,そこで作られたリンパ球の一部は血液に入り,他の一部は遊走細胞となって組織の中を動いてまわる.リンパ性組織とリンパ球との関係は赤色骨髄と赤血球との関係と同じである.リンパ球が赤血球にかわることができるかどうかは疑問である.血小板に相当するものはリンパには存在しない.リンパ漿もまた線維索を生ずることによって凝固する(Fibringerinnung).
3.筋組織Muskelgewebe
筋組織は個々の収縮性要素すなわち筋線維Muskelfasernにより成っている.
筋肉においては原形質の収縮性という特徴が高度に進んでいる.
次の3種が区別される:
1. 横紋筋線維gestreifte(随意性willkurliche, 動物性animale) Muskelfasern. 随意運動のできるあらゆる筋肉およびその他の場所にある(後述を参照のこと).
2. 平滑筋線維glatte8食物性vegetatie) Muskelfasern. 呼吸道,消化管,泌尿生殖道,血管壁,皮膚の中にある.
3. 心筋線維Herzmuskelfasern. 心臓壁の筋層をなし,また肺動脈と肺静脈の壁に多少とも及んでいる.
1. 一つの横紋筋線維は筋鞘Sarkolemm,筋形質Sarkoplasma, 原線維の形をとってならんでいる収縮性の物質,および多数の核より成る.
その形からいうと,横紋筋線維は長く引きのばされた柱状を呈して,その横断面は不規則な円形である.その両端は丸みをもって終わることもあり,円錐状あるいは斜めに切ってその端を鈍くしたような形をしているものある.この筋性にが枝分かれすることは少なくないが,舌の筋肉では枝分かれした横紋筋線維が良く発達している.
その長さは12cmに達するのものがある.太さは30µと70µの間である.同一の筋の中でいろいろ異なる太さの筋線維が隣り合わせて存在する.
一定の筋の中にある筋線維の数に関していえば,良く発達した新生児の肩甲舌骨筋M. omohyoideusは20,808本の筋線維をもつが,弱々しい筋肉の発達を示す男では同名の筋がわずか14, 251本の筋線維しかもっていない(Riedel).
各々の横紋筋線維はうすい透明な膜すなわち筋鞘Sarkolemmで包まれている.この膜は以前の考え方では細胞膜に当たるものであって,核をもっていないのである(図108)比較的近年の研究によれば,この膜は結合組織から生じたもので,格子線維をもっている.
この膜は煮ても溶けず,うすいアルカリや酢酸の中でも溶けない.しかしトリプシンで消化される.この膜は透明であるから,新鮮標本や保存標本では特別な方法を用いたときにのみみることができる.生の筋線維では機械的な作用あるいは浸透圧の影響によって筋線維の内容が断ち切れて,筋鞘は害せられないで残ることが時としておこる.そのときに筋鞘が断ち切れた内容の両端の間を橋渡ししており,あるいは図108右側のように筋鞘が胞状にもち上がっている.また個々の筋線維の切断端において筋鞘が外方にめくれた膜としてみられることがある.
この筋線維の最も目立つ特徴は横紋quere Streifungである.明暗の横の縞が交互に並んでいる.明るい筋の所は光線を通すのであって,個々は単屈折性einfachbrechend (isotrop)であり,暗い筋の所は複屈折性doppeltbrechend (anisotrop)である.横紋は各筋線維を構成している多数の筋原線維Myofibrillen (図107)の特別な性質に基くのである.この原線維は筋形質Sarkoplasma oder Sarkogliaとよばれる筋性にの原形質によって収束されている.この筋形質の中に多数の細長い核がたがいにある距離をもって存在し,これが人間や哺乳動物では筋鞘のすぐしたにある.普通は各々の核の所にやや多量の筋形質が集まっていて,これが色素顆粒を含んでいることもある.あるいは細い顆粒性の筋が核の両端に接してみられる.また核の近くでなく,筋形質の筋の中にも顆粒の列が存在することがあって,これを間質顆粒interstitielle Körnerという.
筋形質によってまとめられて,原線維の多量あるいは少量が1つの束をなしており,またさらに隣接する原線維束の間をへだてて,筋形質の比較的大きい集まりがある.そのために1本の筋線維を横断したときのきれいな像が生ずるのであって,個々の原線維は点としてみえ,その集束は1つの小区画としてあらわれる.これがコーンハイム野Cohnheimsche Felderとよばれるものである(図111).
核の豊富さKernreichtumはすべての筋線維で同じというわけでない.

[図106]横紋筋線維 ×400. 横紋が明瞭にみえる.縦の筋は所によってわずかにみえる程度である集束の縁のところで筋鞘の観の輪郭がみえる.筋線維の核が所々ではっきりと透視できる.

[図107]筋原線維 それをばらばらにほぐした標本.×500.
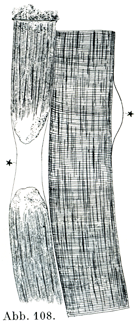
[図108]筋鞘を示す標本 カエルの新鮮な横紋筋線維.細析して水を加えたもの.星印の個所で筋鞘がみえる.
多くの動物例えば家兎では2種の横紋筋がある.赤いのと白いのとで有って,前者の例は半腱様筋,ヒラメ筋であり,後者の例は大内転筋である.赤い筋では横紋が比較的に不規則であり,縦の筋が割合よくみえる.また同時に赤い筋は円みのある核をたくさんもって,その一部は筋線維の深部のもある.電気で刺激すると,赤い筋は白いものよりもゆっくりと収縮する.若干の動物では赤白の各線維が別々の筋をなしているが,他の動物および人間では両種の線維が1つの筋の中にまじって存在している.

[図109]横紋筋線維の横断で核の位置を示す.ヒトの手の虫様筋.×200.
一本の筋線維およびその含む原線維が層をなすことSchichtungについて,図106は単屈折と複屈折の物質から成っていることの簡単な,そして弱拡大で見た様子だけをしめしているにすぎない.適当な材料でさらにくわしくしらべると,この両物質の内部でさらに詳細の点が明らかになる.明るい単屈折のなかに,それをちょうど折半する暗い1線の間膜Krausesche Querlinie(横線の意)によって先般されている.また暗い間膜の両側に副盤Engelmannsche Nebenscheibeがみられることがある.この分かれ方gliederungはもちろん個々の原線維にも成りたつのである.それゆえ,1つの筋要素Muskelelementは順にいうこと次の8部からできている.
1. 副盤Engelmannsche Nebenscheibe
2. 単屈折質isotrope substanz
3. 副屈折質anisotrope Substanz
4. 中膜Hensensche Mittelscheibe
5. 副屈折質
6. 単屈折質
7. 副盤
8. 間膜Krausesche Querlinie
この区分がどんな機能上の意義をもつかについていうと,刺激によって筋線維が収縮するときは,明るい単屈折の部分には入りこむのであって,そのために後者は高さが特にかわらないで,幅が増すのである.原線維はそのために短く,しかし幅広くなる.明るい横縞はかくしてほとんど全くなくなり,暗い横縞がまるで接触するほどたがいに近づくことがある.同じ現象が1本の原線維のかなり広い範囲にわたって起こるし,また1本の筋線維がもつすべての原線維に同じことが起こる.だから1本の筋線維のはたらきが何百万という微小なはたらきの総合である.われわれは1個の筋全体がすでにはなはだ著しい仕事をなしうるものであることを理解するのである.刺激をあたえたことがどうしてこのような物質の移動をおこすのか,収縮がやんだときにどういうぐあいに物資が戻ってゆくのか,確かなことはまだ分かっていない.それはもはや全く分子の問題にはいっているのである.
上のことからして,単一の収縮性の要素としては,原線維の一部で2つの間膜の間にある1から7までの部分であるか,あるいは1つの間膜からそのとなりの中膜までの部分を考えるべきであろう.前者自身が機能的にみて,すでに2重の要素と思われるので,上に述べた第2の可能性が生まれるわけである.
筋線維や原線維を変更でみると,一定の部分が単屈折性,他の部分が複屈折性にあらわれる.最も簡単な場合には複屈折性の暗い盤と単屈折性の明るい盤とが交互に並んでいる.いる.間膜がみえるときはこれも複屈折性であり,副盤は単屈折性である.
筋の化学的検査でまず分かるのは,細胞の一次性分として前に述べたものはすべて筋線維に含まれていることである.その中で蛋白質は大量であり,ヌクレインはごく少量であり,そのほかヒポクサンチン,クサンチン,レチチン,コレステリンもある.無機部ととしてはカリウム,マグネシウム,カルシウム,鉄,燐酸が存在する.二次性分としては沢山のものがある::血液色素(ヘモグロビン),ケラチン類似Keratinoidの物質(これは筋鞘からくる),溶解している酸素類,クレアチン,クレアチニン,カルニンKarnin,グアニンGuanin,尿酸,尿素:タウリンとグリココル:イノシン酸Inosinsäure,プロト酸Protsäure, グリコーゲン,デキストリン,糖類,スチリットScylit,イノシットInosit,乳酸および食塩(A. Kossel).
死に伴って蛋白質の化学的変化が当然起こるので,酸が形成されて,筋(平滑筋もまた)の最終のそして長く継続するつよい収縮が引き起こされる.これがいわゆる死硬直TotenStarreである.
死硬直と同じ現象が個々の筋に,それへの血流が絶たれたときにおこることがある.
前身における筋肉の分布という点では,多核性の横紋筋線維ははなはだ広い分布領域をもつが,単核性のそれは心臓にだけみられることが注意されなければならない.多核性の横紋筋組織はとくにすべての骨格筋の主成分として,また眼球のまわりや聴覚器,消化管の始まりと終わりのところ,呼吸器の初めの部分や泌尿生殖器に存在する.なお注意を要するのは各種の筋組織の分布に関して動物界では大きい差異が存在することである.皮下組織学の教えるところによると,筋肉には個々に述べる以外の型のものもある.
傷ついた筋線維が再生Regenerationをするのは次のごときぐあいである.筋線維の若干の部分がまずもって壊れてしまうと,筋形質とその中の核とが盛んな活動をはじめて,両者とも増加する.ついで新しく生じた原形質海のなかにふたたび原線維の分化があらわれてくる.しかしいっそう大きい物質欠損の場合には結合組織性の瘢痕が作られる(Sokolow).
横紋筋線維における神経終末については筋学を参照のこと.
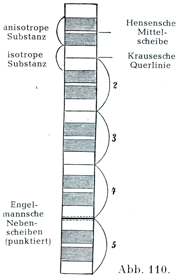
[図110]1本の筋原線維の区分を示す模型図 2~5:それぞれ1個の筋要素.
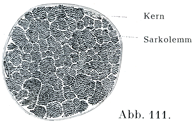
[図111]コーンハイム野 横紋筋線維1本の横断.(人の口蓋帆拳筋.)×1000.

[図112]中くらいの太さの横紋筋線維(人)の一部(Sharpeyによる,Quainの本からの引用).間膜Krausesche querlinieがみえる.×800.
2. 平滑筋線維glatte Muskelfasernは多くは紡錘状の細胞(図113)であって,人ではその長さが50~225µ,幅は4~6µである.平滑筋細胞尾発見者であるKollikerはその長さの極小,極大として,22µと560µをあげている.細胞膜は欠けていて,そのところに原形質の辺縁の1層がある.細胞の縁は平滑であるが,たぶんここに細い溝が刻まれているのであろう.細胞体は多くの場合に密集した縦の細条すなわち原線維を,その明瞭度はちがうが示しており,この細条は細胞体がその両端に向かって細くなるにつれて,だんだんと終わってしまう.横断面ではこの原線維は密集した点としてみえる.原線維の間にある物質は筋形質Sarkoplasmaである.
細胞の中央部に長い卵円形,あるいは棒状の核が1個ある.比較的長い線維だと核が2つあるいは3つある.核の両端に接して細かい顆粒をもつ原形質が少しく集まっている.核は多少の差はあるが概して明瞭なクロマチン網を有し,核小体は2個あるいはそれ以上の数に存在する.核が縦に伸びている面のそばに原形室内に双心子(中心小体)がある.
多数の筋細胞がたがいに密集して,かなり大きい束や膜をなすところでは,個々の筋細胞は厚されて平たくなり,多面をもつ稜柱状となる.
それは横断してみたとき筋細胞が大小いろいろの,時にははなはだ規則正しい多角形の部分としてみえることから分かるのである.筋細胞の間に結合組織があって,これが個々の筋細胞を包み,それらを一と纏めにすると同時に個々の間をへだてているのである.比較的大きい隙間のあるところは,それだけ結合組織が多く集まり,血管,神経などがそこを通っている.
紡錘状の平滑筋線維の他に,枝分かれする細胞体をもつものがある(図114).後者はすでにKöllikerやその他の人々によって記載されてきた.比較的新しいところでは,Benninghoff (Z. Zellforsch., 4., Bd.,1926)が心臓内膜において,Stieve(Z. mikr.-anat. Forsch.,17. Bd.,1929)が子宮筋層において,枝分かれする細胞体をもつ平滑筋線維をはなはだ明瞭にあらわし示したのである.Benninghoffによれば,紡錘状の平滑筋線維はいずれの点からみても発達の終わりの段階にあるもので,沢山の枝分かれをしている平滑筋線維は発達の初めの段階にあるものとみなされるのである.そのあいだの数多くの段階のものがあって,紡錘形と分子型は移行する.
変更でしらべると,平滑筋細胞は1軸性に複屈折が陽性である.その軸は細胞の長軸に平行している.
平滑筋は体の中では多量に存在するのであって,あるいは密な膜の形をなし,その膜の重なり合った各層がたがいに違った方向,それも多くの場合たがいに直角をなす方向に走る筋線維の集まりであることが普通である.あるいは小さい梁や網,もしくはむしろ密な塊というような形を呈している.
汗腺(毬状腺)では外胚葉性の上皮細胞,気管支の細い枝primitive Bronchenでは内胚葉性の上皮細胞が平滑筋細胞の起こりをなしているようである(筋上皮細胞Epithelmuskelzellen) (Kölliker, Stieda).眼の瞳孔括約筋M. sphincter pupillae (Nussbaum)および瞳孔散大筋M. dilatator pupillaeも同様に外胚葉性のものである.
平滑筋細胞が数を増すことVermehrungは有糸分裂によっておこる.そのさい核分裂が必ずしも常に細胞体の分裂を伴わないので,多核の平滑筋細胞ができる.傷によって生じた平滑筋の欠損もまたこれと同じ行き方で補充される.
平滑筋の神経終末については第II巻の交感神経の項を参照のこと.
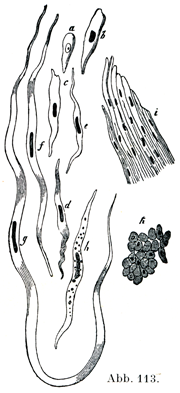
[図113]ヒトおよび哺乳動物の平滑筋線維.a 2インチのブタ胎児の胃壁より得た平滑筋芽細胞;b その発達がさらに進んだもの;c~g ヒトの平滑筋線維の種々な形;h 脂肪顆粒を有するもの;i 平滑筋線維の集まった1束;k ウシの大動脈壁の平滑筋束の横断,多くの核が切断面にみられる.(Freyによる.)
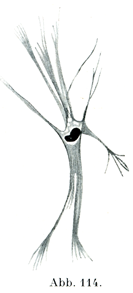
[図114]枝分かれしている平滑筋細胞 ヒトの右心室の内膜より.×400. (Benninghoff, Z. Zellforsch. 4. Bd.,1926)
3. 心筋線維Herzmuskelfasern oder Herzmuskelzellenは短い円柱状の細胞で,横紋を示し,また非常に細かい格子線維を含む筋鞘を有している.核は1~2個で,細胞の中軸にあり,この点が骨格筋では核が筋鞘に密接しているのと違っている.心筋線維の横紋は体幹の横紋筋のにくらべてずっと繊細である.また縦の方向にも細かい筋がついているのは原線維よりなることを示すのである.この細胞の両端をなす面は段階状に切りこまれていて,しかしその面は全体としてみると線維の縦軸に対して横走している.側方に出る突起がやはり階段状(テラス状)にぎざぎざとした面をもって,隣りの細胞あるいはその突起と合している.細胞がたがいに合するところに特別な物質(接合質Kittsubstanz)の薄い1層がはさまっている.
心筋線維はたくさんの枝分かれと合同をするので,全体として筋肉性の網状物(叢Plexus)をなし,その網の目ははなはだ鋭い角をしている.楕円に近い形の核が細胞の中軸にあって,そのまわりをとりまいて比較的多量の筋形質が紡錘状をなして集まっていて,そこにすでに若い人でも黄褐色の色素顆粒がみられる.細胞体は筋形質に包まれた横紋性の原線維と同じなのである.しかしここで次のことを述べておこう.心筋線維では原線維の集束が一部は稜柱状であり,一部は帯状をしていて,その帯状の束は細胞の周辺にあって,細胞の中軸に対して放線状の配列を示しており,稜柱状のものは細胞の中軸の近くにあって,帯状の束によって取り囲まれる状態である.
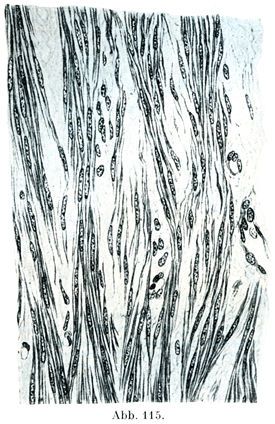
[図115]平滑筋の縦断17才の女の子宮筋層.×400. (Stieve, Z. mikr.-anat. Forsch.17. Bd.,1929. )
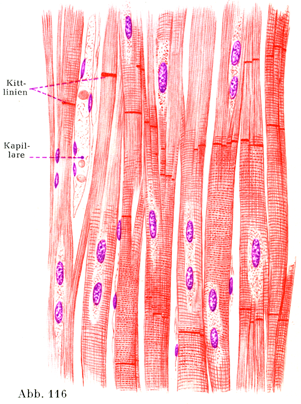
[図116]ヒトの心筋の縦断
接合腺Kittlinienは場合によって,いろいろと異なる量にみられ,またはなはだ不規則な配置であらわれる.時としてそれがみられぬことがある.かなり多くの学者は心筋が個々の心筋細胞から組み立てられていることを否定している.できあがった心臓では全部の筋肉が単一の立体的網状を呈する合胞体であって,その個々の分節が介在部Schaltstuckeすなわちここにいう接合線によってたがいに結合している.この介在部は光輝線Glanzstreifenともよばれ,比較的新しい考え方によると心筋線維そのものに属しているのであって,そのことは心筋が収縮したときこの線が単屈折性となり,心筋が弛緩したとき,これが複屈折性となることから分かるという.しかしK. W. Zimmermannはその2人の女弟子,PalczewskaおよびWernerとともに新しい説をしりぞけて,古い方の見地が正しいとしている.心筋の起原は臓側板Splanchnopleura(中胚葉の側板の一部で内臓表面を被う部分)にある.
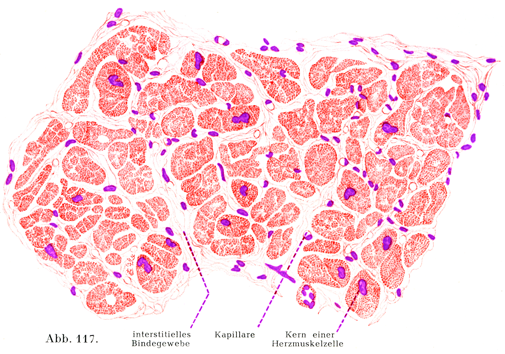
[図117]ヒトの心筋の横断
4.神経組織Nervengewebe
神経組織は2種の形態要素からなっている.その1つは神経性のものでニューロン(神経原,神経単位)Neuronen,他は支持細胞でグリア細胞(神経膠細胞)ellenあるいは短くGliazellen (R. Virchow)である.
まずニューロンについて述べる.
ニューロンとは1個の神経細胞とこれが有するすべての突起をその末端まで合わせたものである(Waldeyer).
神経細胞Nervenzelle(あるいはあまり結構でない言葉であるが,Ganglienzelleともいう)とはニューロンの細胞体の部分とその突起の始まりのところを指すのである.
神経細胞は主として中枢神経系のなかにあるが,また脳脊髄神経および交感神経の経過中にも豊富にあり,体の全くの末梢部にも存在する.
細胞体からでる突起に2種が区別される.
a)樹状突起Dendriten(原形質突起Protoplasmaforstätze).これはしばしば数が多くて,その名が示すごとく,豊富に枝分かれをし,それも多くはほぼ同じ太さに2分するのであって(dichotomisch),細胞体のわりに近いところで終わる.
b)神経突起Neurit, Nervenfortsatz(軸索突起Achsenzylinderfortsatz)はたいていはただ1本だけあって終わるが(図119),しかし多くの場合長い距離を走った後に,その終止する場所に達する.
この突起は中枢器官の中をとおる間に細い側枝Kollateralenを多くは直角をなしえ出し,側枝はまもなくごく細い枝でできた叢すなわち終末分枝Endbäumchen, Telodendronをもって終わる.軸索突起もまた1度あるいはそれ以上の回数,2つの同価置の枝に叉状dichotomischに分かれることがある.それは図118の運動性ニューロンの模型図に示されている.
A. まずわえわれはニューロンの細胞体の部分すなわち神経細胞をみよう.これは細胞膜をもたず,細胞の形と大きさはいろいろである.球形のもの,平たいもの,紡錘形のものがある.それからでる突起によってしばしば星状,錐体状などの神経細胞がある.大きさに概して著しいのであって,大きい神経細胞は肉眼でもみることができるほどである.しかし神経系の数多くの場所にははなはだ小さい神経細胞も存在するのであって,その細胞体は簡単な染め方ではほとんどあらわし得ないのである.そこで神経細胞の大きさは10~150µという大きい幅の中で変動することになる.
神経細胞のだす突起はその細胞体の一部に他ならないのであるから,これをも細胞の大きさに加えるならば,多数の神経細胞ははなはだ大きいものとなるのであって,人間では1メートルやそれ以上の長さのものがあり得る.
でてゆく突起の数によって,神経細胞には単極,双極,多極のものが区別される.
突起を一つも有たぬものは,人工的に突起が除かれたものは別として,神経細胞の幼若形である.どの神経細胞でも発達の初期には突起のない時代があった(もちろんそのときのも細胞間橋はおそらく存在したのであろうが,これは問題のそとにおく).単極unipolarの神経細胞すなわち1本の突起をもつものは多数に存在する.しかしこの1本の突起は早かれ遅かれ,ある経過ののちに分岐する.その分かれた枝の1つが神経突起の意味をもつのである.双極bipolarの細胞も同様にたくさん見られる.本の突起は細胞のそう反する場所から出ている.それよりも多数の突起が細胞体から発するならば,多極multipolarの神経細胞である.
樹状突起は広い起始面をもつ細胞体からでる.それは次第に細くなるが,細その叉状分岐までは細胞体じしんと同じ構造をたもっている.神経突起は縦走する細かいすじをもったいわゆる起始円錐Ursprungskegelをもって発する.この突起は細胞のすぐ近くですでに細くなっており,そして初めのうちは分岐や側枝を示さないのである.
細胞体は新鮮なままでとり出した神経細胞では,どこも個々も同じように顆粒が密に含まれて濁って見える.特別な包蔵物としては黄色ないし暗褐の色素と脂肪の小滴がたやすく認めあれる.
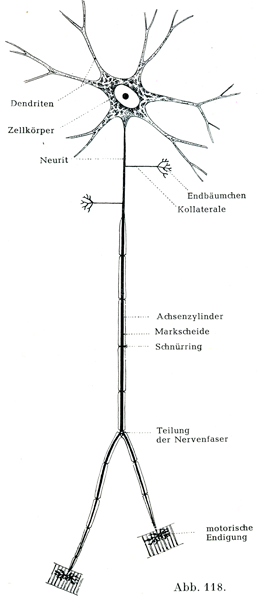
[図118]1個のニューロンを模型的に示す. 脊髄の前柱の運動性神経細胞の構造をもとにして.
神経細胞の色素含有は個体の年齢とともに増す.しかしまた中枢神経の定まった場所にははなはだしく色素に富む多数の神経細胞が集まっている.それは例えば青斑Locus caeruleus,黒核Nucleus nigerである.
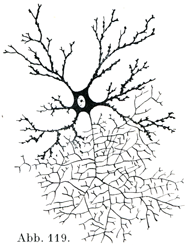
[図119]細胞体の近くで枝分かれして終わる神経突起をもつ神経細胞.(Schäferによる.)
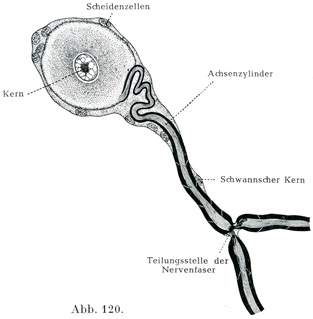
[図120]単極神経細胞 家兎の三叉神経の半月神経節より.(KeyとRetzius. )

[図121]双極神経細胞 ダツ(硬骨魚類)の三叉神経節より.(Bidder. )
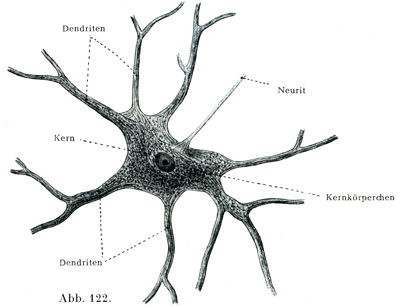
[図122]多極神経細胞 ウシの脊髄灰白質の前柱よりとりだしたもの.×250.
神経細胞にとってとくに特徴的なのはその大きい胞状の核である.この球または楕円に近い形で,はっきりした薄い核膜と1個あるいはそれ以上の数の核小体をもっている(図14,15,118~124).適当な処理によって初めて繊細な核材がそこにはっきり認められ,新鮮なままの標本ではそれがみられない.この核材はリニンLininのみからなるごとくみえる.
細胞体の内部には次のようないくつかの特別の構造がある.
a)色素好性物質chromophile Substanz (Nissl),
b)神経原線維Neurofibrillen (Bethe, Cajal),
c)内網装置Binnengerüst (Kopsch), Appaato reticolare interno (Golgi),
d)細胞内細管intrazellulare Kanalchen (Holmgren),
e)中心小体zentralkörperchen
a)色素好性物質は普通はニッスル小体Nisslsche Körperchenともよばれ,大小種々の顆粒や細糸がいろいろな形の群をしたものとして存在する.不規則な形で角のある塊があって,その間に狭い明るい場所がはさまれている.その外観はいろいろな種類の細胞で異なっているが,おのおのの細胞種によっては特色がある.細胞の機能状態のによってもちがうので,ニッスル小体を見ることによってニューロンの栄養ないし健康状態をある程度測ることができる.神経突起が切断されたり,神経細胞があまり永くつづけて働いたりすると,ニッスル小体が減少し,また消失することがある.その他の変わり方としては濃縮した塊をなすこともある(図14,15).
b)神経原線維はすでに古くM. Schultze, Kupffer等がみたものであって,はなはだ細い線状のものとして細胞体の中のみならず,樹状突起および神経突起の中にも見られる.原線維は細胞体の中でニッスル小体のあいだの明るい場所を通っている.原線維が細胞体の中および樹状突起の幹の中にどんな具合になっているか,その詳細の点がしばしば議論されて,しかもなお決定していない.それについては,原線維じしん,あるいはその集束がたがいに直接につづいているのか,もしくはこれらは独立しており,ただ隣り合わせて走っているのかということが議論の種になっている.Betheによると原線維は網をつくらないで,細胞を貫いて走るのが普通なのであって,格子状の構造をなすことはいっそう稀である(これは無脊椎動物に見られ,脊椎動物の脊髄神経節の細胞,シビレエイTorpedo marmorataの電気葉Lobus electricusにもみられる).この説に反してCajalやDonaggioその他の人々は原線維が細胞体の中でも樹状突起の最初の分岐角のところで網状の配列をしていることを断乎として唱えている.最もこの人たちも原線維束の一部が他のものと合したりしないで細胞を貫いてとおることも認めてはいる.また樹状突起や神経突起の中では原線維が束をなして,たがいに隣りあって存在している.
c)内網装置(図124) (GolgiのApparato reticolare interno)はいろいろの太さの円るい形の索状物が集まってはなはだきれいな網をしているのであって,この索状物が時としては顆粒からできているようにみえることもある.これは細胞体の内部にあって,その表面には達してない.網の一部が核に触れていることもあるが,核の近くでも,また細胞体の辺縁のところでも狭い一体が内網装置を全くもっていないことが普通である.その上に網をなしている糸が側方に枝を出して,まもなくこの枝が単に円く,あるいは頭がふくれた混紡の形で終わったり,また(比較的年配のもので)網の若干の部分が小葉状の配置をとったりするので,この装置の像はますます複雑となる.内網装置の本体と意義については11頁ですでに述べた.
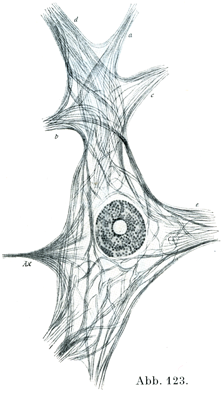
[図123]神経原線維 18才の男の腰髄の大きい前柱細胞.AX神経細胞,a, b. c. dなどはみな樹状突起.原線維はその一部のみが描かれてある.(A. Bethe,1900)
d)神経細胞における細胞内細管intrazellulaare Kanälchenは特別な方法によってあらわすことができる.現在のところ分かっているのは,神経細胞の細胞体の内部に長短いろいろの細い管でできた1系統があって,この間は固有の種をもたず,また細胞の外にも開いていないので,細胞体の中を貫いているということである(図124).
Holmgrenによるところの細胞内細管は細胞の表面に開いているという.彼の意見によれば神経細胞の細胞体は他の種類の細胞が出す固い突起の集まった網状をなすものによって貫かれている.この網状物はその機能の上から栄養海綿体Trophospongiumとよばれる.この網の諸部分が溶けることによって細胞内細管が生ずるというのである.
e)中心小体Zentralkörperchenはいろいろな種類の神経細胞において,数多くの動物および人間でも見られている.
B. 樹状突起Dendritenは原形質突起Protoplasmafortsätzeともいうが,豊富にまた細かく枝分かれするので,この突起はすべての神経細胞にあるとは限らない.例えば脊髄神経節の細胞には欠けている.これが細胞体から発するところは幅がひろくて,そこから漸次的に細くなる.この点は神経突起との差異として特記すべきものである.樹状突起の分岐はたいてい同じ太さの二た叉におこる.非常に細くなった枝が中枢神経系の汎在基礎網allgemeines Grundnetzに終わっている.
樹状突起およびその枝の全部は神経原線維とその間にある原線維間物質あるいは原線維周囲物質interoder perifibrillare Substanzから成っている.その主な幹の中にはニッスル小体が最初の叉状分岐のところまで細胞体の方からつづいており,時としてそこを越えてなおいくらか達している.叉状に分かれる角のところにしばしば色素好性物質の三角形の塊がある.またこの場所でCajalは神経原線維が1つの側枝から他の側枝に直接に移行することを見ている.こういう原線維は細胞体じしんとは何らかの関係をも持たないわけである.樹状突起の比較的太い枝は原線維の束をもっているが,最も細い枝はわずか1本の原線維とそのまわりのいくらかの原線維周囲物質とから成っている.神経細胞が死んで,それが保存されたばあいに,原線維周囲物質がいわゆる静脈瘤に似た状態Varikositätenをなす.少数の個所にそれが集合して,その他のところでは少なくなり,かくして真珠を連ねたような像が生ずるのである.
C. 神経突起Neuritは軸索突起Achsenzylinderfortsatz (Axon, Neuraxon)ともいい,すでに述べたように,多くはただ1本存在する.これは幅のひろい起始円錐Ursprungskegelをもって細胞体からおこり,急に細くなって,ついで細胞体からかなり離れたところで漸次的にふたたび太くなる.時として神経突起が細胞体からでなくて,樹状突起の幹からでている.また神経突起が多極神経細胞には常に存在するとはかぎらないで,全く欠けていることがあり,また2本以上存在することもある.
起始円錐はニッスル小体をもっていないので明るくみえる.そして他の方にむかって集中する細かい縦のすじを示し,そのすじの間にばらばらに離れた顆粒が列をなしている.この縦のすじは細胞体から神経突起の中に入ってゆく原線維のあらわれである.原線維はBetheがいうように,その細胞がもつ全部の樹状突起からやってきたものである.しかしおのおのの樹状突起が導いている原線維の量は決してこの樹状突起の太さやその力強さに比例してはいない.
神経突起の中に集まった原線維はまずたがいにごく密接になるのである.そして細胞体からいくらか離れたところで初めて比較的多くの原線維間物質がそのあいだにはいることによって原線維の密接状態が解けてほぐれるのである.
固定して保存した標本では原線維が限外顕微鏡的な太さの現々線維Protofibrillenの集合によってできている.(Baud, C. A., Acta anat.10. Bd.,1950. )
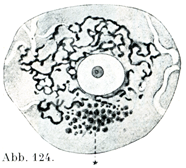
[図124]内網装置 脊髄神経節の小さい神経細胞.Kopsch-Kolatschevの方法,同時にHolmgrenの細管の数個が示されている.×1000. *は色素.
それと同時に多くのばあいは(例外:嗅糸Fila olfactoria)細胞体からいくらか離れたところで,神経突起を周囲から隔離するための被いが初めてあわれる.この鞘の種類によって有髄と無髄神経線維markhaltige und marklose Nervenfasernが区別される.
有髄神経線維の構造としては1本のガラス管に中に幾本かの細い糸を通して,且つ水をもって充たしたものを考えると,かなり真に近いのである.管のガラス壁が髄鞘に当たり,髄鞘でかこまれた内容すなわち軸索は上に述べた模型の細い糸と水にそれぞれ相当する神経原線維と原線維間物質から成っている.
軸索Achsenzylinderは新鮮な標本では灰白色で,太さは0.5~1.0µである.髄鞘は輝いてみえる.末梢神経では髄鞘のまわりを神経鞘Neurilemm (Neurolemmというのがいっそう宜しい)がかこんでいる.
したがって有髄神経線維がさらにシュワン鞘Schwannsche Scheideをもつものと,これをもたないものとに分類される.
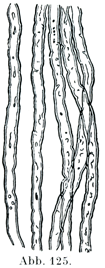
[図125]新鮮な有髄神経線維 重複した輪郭をもち,またでこぼこした境界を示している.(Quainより.)
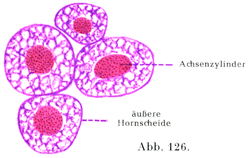
[図126]有髄神経線維の神経角質材.縦断.×1200.
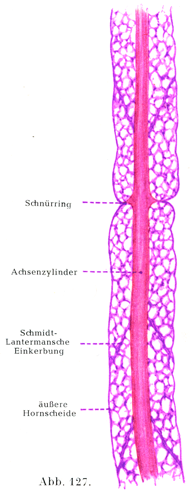
[図127]1本の有髄神経線維の神経角質材.縦断.×1200.
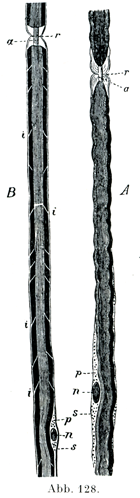
[図128AとB]オスミウム酸で処理した有髄神経線維.a軸索;sシュワン鞘;n, n神経角(髄鞘を軽く凹ませている);p, p角の両極に接している微細顆粒性の物質;r, rランヴィエ絞輪(ここでは髄鞘がなくなって,軸索がみえている);Bにおけるi, iはシュミット・ランターマン切痕に相当する円柱円錐節zylindrokonische Segmenteの境界線である.
a)シュワン鞘をもつ有髄神経線維が脳および脊髄からでるすべての末梢神経の最も主な成分をなすが,なおその他に交感神経系にもある.
髄鞘Markscheide, MyelinscheideはミエリンMyelin, Nervenmarkとよばれるつよく光線を屈折して輝いてみえる物質から成っている.これは蛋白質とリポイドの混合物であって,その上になおニューロンケラチンNeurokeratinという角質様の物質を含んでいる.ニューロンケラチンが溶液の形で存在するのか,あるいは髄鞘の中で網状の構造をしているのかは定まっていない.新鮮な神経線維では(図125),髄鞘ははっきりした境界線を示し,髄鞘の存在する範囲ではその中に包まれている軸索を外から見ることができない.しかし死についてある神経線維だと,まもなく第2の,すなわち内方の境界線があらわれてくる.かくしてその線維は複輪郭性doppelt konturiertにみえる.この複輪郭性はそれにつづいて高度におこるところのミエリンの凝固現象の最初のあらわれである.神経にアルコールとエーテルを加えて煮て,ミエリンから脂肪性の部分を除き去ると,それまでミエリンで充たされていた場所にきれいな網状の構造が残るのであって,これが前にも述べたことのある髄鞘の神経角質材Horngerüst, Hornspongiosaである.この構造は染色することによって,例えば鉄ヘマトキシリン(Corning)やその他の方法で切片標本でもみることができる(図126,127).
髄質の内側と外側の境界をあらわすところの,同一物質より成る2枚の鞘(外および内角質髄äußere und innere Hornscheideとよばれる)があって,その間に多数の小さい梁が張られており,この梁と鞘を合わせたものが神経角質材である.
シュワン鞘は神経鞘Neurilemmともいい,末梢神経線維がもつグリア性の物質であって,ガラスのように明るくて,弾性をもち,無構造にみえる膜で,髄鞘の外面に密接している.そしてシュワン鞘の内面に接して,ある間隔をおいて楕円に近い形の核が存在する.その核のまわりに多くのばあい,細かい顆粒を含む原形質があって,これが特に核の両極のところで集まっている(図128).
髄鞘にはその他になお特異な切れ込みがある.すなわちランヴィエ絞輪Ranviersche Schnürringeとシュミット・ランターマン切痕Schmidt-Lantermansche Einkerbungenとである.
ランヴィエ絞輪ではシュワン鞘もまたその形成に大いに与っている.シュワン鞘の深い輪状の絞れが,ある間隔をおいて軸索のすぐ近くまではいりこむので,ここでは髄質が全く欠けていることがある.軸索は絞輪のところで円錐状に太くなっていることがある(図127,129).硝酸銀で処理したばあいに,そのへこみの奥に黒く染まった横走の境界線があらわれる.それは上皮の接合線と似ている.それゆえシュワン鞘は管状をなす分節の集まりでできているのだろうと考えるのである.銀液はここからその程度は個々のばあいによって違うが,多少とも軸索の中に或る距離だけ及ぶのであって,かくしてラテン十字lateinisches Kreuz (Ranvier)[銀十字Silberkreuzと同意語である.(小川鼎三)]とよばれる像が生ずるのである.銀の沈着した軸索がしばしば横の縞模様を呈して,明るいすじと暗いすじが交互に重なり合っている.これがフロムマンの横線Frommannsche Linienである.神経線維じしんは絞輪によって外面的にはいわゆる輪間節interanuläre Segmenteに分かたれている.輪間節の長さすなわち絞輪間の距たりは細い線維で0.08mm,太い線維では1mmに達する末梢の有髄神経線維のすべてが絞輪をもっている.神経根の線維も同じである.
絞輪の役目としては軸索と周囲のリンパとのあいだの物質の交流をたやすくする.かくしてその存在は軸索の物質代謝に役立つと考えるべきであろう.
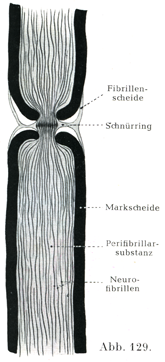
[図129]軸索の構造 (Betheによる.)
切れ込みの第2の型は髄鞘のみに関係するのであって,1個の輪間節の中で,髄鞘はシュミット・ランターマン切痕という細かい漏斗状の切れ込みによって,1列をなしてならぶいくつかの小さい円柱円錐分節zylindrokonische Segmenteに分かれている.シュミット・ランターマン切痕はあるいは中心端の方へ,あるいは末梢端の方へ傾いた斜めの方向をとっている.そしてすでに新鮮な神経線維でも見られるのである.この切痕をつくっている詳細な物質は接合質に近縁のものである.
b)シュワン鞘をもたない有髄線維は多量に集まって中枢神経や視神経の白質を形成している.シュワン鞘がない変わりにここには中枢性のグリア(神経膠)zentrale Neuroglia(図139)があって,その配列の具合といっそう微妙なはたらきにより末梢性のグリアであるシュワン鞘の細胞Schwannsche Zellenとははなはだちがうものといえる.
2. 無髄神経線維marklose Nervenfasernは特別な被いを全然もたないか,もしくはシュワン鞘のみをもっている.もっともシュワン鞘の核Schwansce Kerneがこれに伴っている.
a)シュワン鞘をもつ無髄神経線維はまた灰色線維graue Fasern, または膠様線維gelatinöse Fasernとよばれる.主として交感神経にあるが,また豊富に末梢神経の中に存在する.これは非常に薄い髄鞘をもつので,髄の少ない神経線維markarme Nervenfasernとよぶのがいっそう正しい.
b)シュワン鞘を欠く無髄線維は裸軸索nackte Achsenzylinderという不当な別名をもらっていたことがあるが,そのかなり長いものは人間では嗅糸Fila olfactoriaにあるのみである.これは嗅上皮存在する感覚細胞の中心性突起であって,0.5µの太さにも達しない.
初めから無髄の線維,あるいは途中で無髄になった線維が末梢部で,そのシュワン鞘を失うと,軸索あるいは原線維束が裸になるわけである.末梢の末梢部はこの点では脳および脊髄で神経線維がおこるところと似ている.そのおこる所は髄鞘をもたず,またシュワン鞘もないのである.シュワン鞘をもたない無髄の神経原線維や原線維束は身体の末梢部のみでなく,中枢神経の内部のも求心性に導く神経の終末分枝の所に非常にたくさん存在している(AmbronnとHelkd, Sitzber. sachs. Ges. Wiss., Leipzig ,1895).
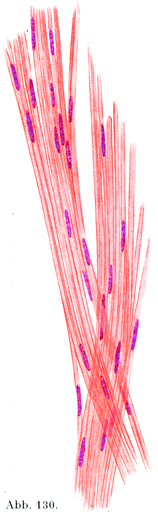
[図130]無髄神経線維 解いてばらばらにしてある.
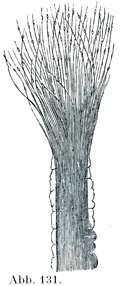
[図131]ダツ(硬骨魚類)の鼻窩より得た神経原線維束.ほぐれてごく細かい静脈粒状の原線維となっている.(M. Schultze)
神経線維の太さKaliber der Nervenfasern
神経原線維は中枢性のものと末梢性のものとを間わず,すべてほぼ同じ太さである.しかし原線維の集束すなわち軸索ははなはだしく太さが変動する.
神経線維が全体としてやはり著しい太さの差を示す(人間で1~16µ).太い神経線維は概して太い軸索をもっている.知覚神経と運動神経の一定した目印となるようなことは太さの点では存在しないが,それでも知覚神経の方が個々の線維の太さでいっそう大きい変動をあらわしている.しかしKirsche(1952)によると,運動神経と知覚神経で軸索の直径を比較すると,計測できるだけの差異が“平均において”みとめられるという.神経線維の長いもの,また上方でおこるもの,そしてまたおそらくたびたび使う神経線維は太くなっているようである(Schiwalbe);しかしなんといってもまず豊富な終末分枝や豊富な結合をもつことが太さの増加にあずかっている(Cajal).動物の体の大きさとその神経線維の太さのあいだに直接の相関関係にくらべると割合に細い神経線維をもっているようである(Hägguist, Anat, Anz.,96. Bd. ).
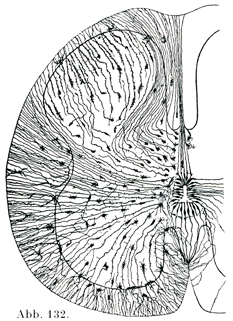
[図132]上衣細胞とグリア細胞 14cm長さのヒト胎児の脊髄.(v. Lenhossek. )クローム銀染色法.
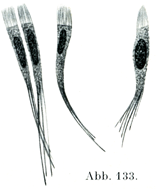
[図133]上衣細胞と上衣線維 家兎の脊髄より.×100. 上衣線維は細胞に近い部分のみが示されている.
神経線維の分岐Teilungen der Nervenfasern
神経線維の分岐ははなはだ多くみられるものであって,それは線維が中枢神経の中をとおるばあいにも側枝などを出すことによって,また末梢では例えば表皮の上皮細胞間隙や筋線維の運動終板に神経が終わることところ,および運動終板に達する少し前のところなどでたくさんみられる.しかし普通に通っている神経幹の内部では分岐は比較的まれにしかおこらない.それに反して脊髄神経節の領域では分岐は当たり前の現象である.末梢の神経線維が分岐するところには常にランヴィエ絞輪がある,図118,120.
中枢性のグリア(神経膠)zentrale Neuroglia.
中枢神経系の支持成分であるグリア細胞(神経膠細胞)には3群が区別される.
a)上衣細胞Ependymzellen,
b)星状膠細胞Astrocyten (蜘蛛細胞Spinnezellenともよばれる),
c)特殊な形のものbesondere Formen.
a)上衣細胞(図132,133).この細胞が集まって上衣Ependymをなしている.上衣は単層絨毛上皮の形で,脊髄の中心管や脳室の内面を被うのである.
この上衣の細胞は楕円円形の核をもつ円柱状の細胞体と長い上衣線維Ependymafaserとから成っている.細胞体の自由表面に小皮縁Kutikularsaumがあって,ここに絨毛が生えており(図133),この毛が運動をすることをLeydingが動物において,Virchow, Kölliker等がヒトにおいて観察したものである.この細胞の一部は分泌機能を持っている(Fuchs, Bargmann).
上衣線維は脳物質の中に遠くはいりこんでいる.胎児の脊髄ではそれが外表面まで達して,そこで膠小足Gliafußという小さい広がりをして終わっている.成人では後正中中隔Septum medianum dorsaleをなしている上衣線維のみが脊髄の外面まで達する.おのおのの線維がいくつかの小線維からできていて,この小線維が早晩たがいに離れるのである.
b)星状膠細胞:すでにごく早い時期の胎児において上衣細胞がからグリア細胞(神経膠細胞)になるものが分かれる.これは上皮性の結合状態から分離して,神経細胞や神経線維のあいだに達する.
グリア細胞に大きな2つの種類がある.大膠細胞Makrogliazellenと小膠細胞Mikrogliazellenであって,その各々がまた2つの種類に分かたれる.前者が長突起細胞Langstrahlerと短突起細胞Kurzstrahler,後者が稀突起膠細胞Oligodendrogliazellenとオルテガ細胞Hortegazellenである.
多くのばあい,オルテガ細胞のみが小膠細胞とよばれる.しかしオルテガ細胞は稀突起膠細胞よりも大きいのであるから,本編のごとくこの2つを合わせて共同の群となして,それを小膠細胞とよぶのがいっそう良い.
大膠細胞(図136 gl)は神経細胞にくらべるといっそう小さい(球形の)核といっそう少ない原形質をもつが,小膠細胞(図136 oglとhgl)にくらべると核が比較的大きくて,原形質もわりあい多い.そして多数の突起をもっているために星状膠細胞Astrocytenとよばれてきたのである.
突起の長短によって長突起細胞と短突起細胞に分かたれる.前者の突起は長くて滑らかで,細くて枝分かれが少ないが,後者の突起は短くて,繰りかえし二た叉に分かれるのである(図134,135).
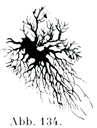
[図134]短突起細胞 ヒトの大脳皮質より.(Golgi原法のKopsch変法による.)
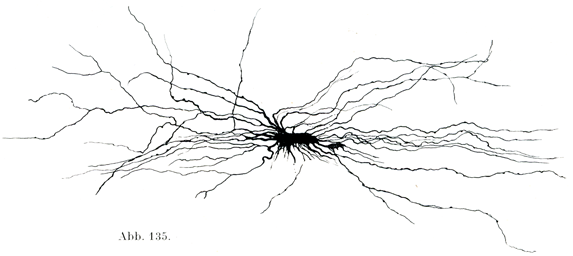
[図135]長突起細胞 ヒトの視神経交叉より.(Golgi原法のKopsch変法による.)
長突起細胞の原形質はまん円い切り口を示す固い線維すなわちグリア線維(神経膠線維)をもっている.それゆえ線維性グリアFaserige Gliaともよばれる.グリア線維は細胞の突起の中にあり,細胞体を貫いて(図138,139),細胞体の表面でしばしばわなの形をして折れまがる.これは細胞の原形質内に生じたものであるが,一部はその母胎である細胞から分かれるために,遊離したグリア線維ができて,これが細胞に属しない特別の構造物とされることがある.グリア細胞はその突起をもってたがいに連続して合胞体Syncytiumをなしている.
短突起細胞(図134,140)はグリア線維を欠くが,原形質内にごく細かい微小粒体Mikrosomenであるグリア粒体Gliosomenをもっている.その突起がさかんに二た叉に分かれて,その先端が広がっていわゆる膠小足Gliafüßeをなして,神経細胞や血管と関係を持つのである.
小膠細胞はいっそう小さいものであって,グリア線維を欠いている.
稀突起膠細胞Oligodendrogliazellenの核は小さくて球形を呈し,多くのばあいクロマチンに富んでいる.その大きさはほぼリンパ球の核にそれに近い.原形質は少量であって,薄い被いをなして核を包んでいる.(図136 ogl).この細胞の突起は細くて短く,枝分かれがごく少ない.原形質はグリア粒体をもっている.
オルテガ細胞Hortegazellenはいま述べた細胞の種類よりは大きい.その核は長めであって,不規則な形である.細胞体は薄い層をなして核をかこみ,突起の数は少ないが,それが豊富な枝分かれをしている(図136 hgl).グリア粒体は存在しない.この細胞の突起は血管壁に付着していない.
HortegaとCajalはオルテガ細胞が中胚葉に由来するものであると考えた(中膠細胞Mesoglia).これははなはだよく運動する遊走細胞である.しかしSpatzはこの細胞を外胚葉性のグリアの特別な一型としている.
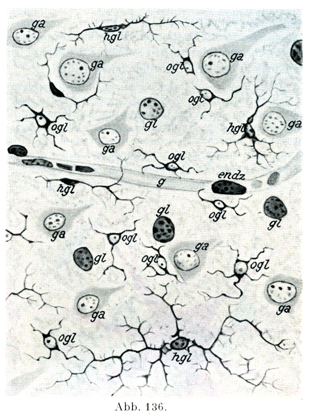
[図136]稀突起膠細胞(ogl)とオルテガ細胞(hgl).ヒトの大脳皮質より.(Handbuch der Psychiatrie. A. I,1.1927. )endz内皮細胞の核;g血管;ga神経細胞;gl大膠細胞の核.
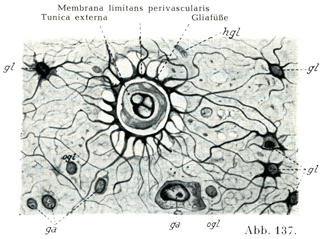
[図137]膠小足と血管周囲境界膜 ヒトの大脳皮質でグリアが増殖しているもの.(Handbuch d. Psychiatrie. A. I,1.1927. )ga, gl, hgl, oglは図136のと同じ.
グリアは中枢神経系の外表面にそって,また中枢神経系の内部の血管のまわりに密な境界膜をなしていて,この膜が神経膠の網および膠小足ととつづいている.
表面境界膜Membrana limitans superficialisは柔膜にぴったりと着いていて,,たぶんこれと固く融合しさえしている.血管周囲境界膜Membrana limitans perivascularisは血管の外膜にすぐ接している.この膜と外膜との間にみられる隙間は人工的産物とおもわれるのである(図137).
c)グリア細胞の特殊な形のものは網膜にあるミュルレル線維Müllersche Stüzfasern,なお小脳皮質の灰白質(Buschelzellen-総状細胞の意)や大脳皮質の表在層にみられるものである(第II巻を参照).
神経組織の変性Degenerationと再生Regenerationについては多数の研究がなされている.いつも末梢の神経幹では編成する線維があるかと思えば,そのそばに再生しつつある線維が存在する.1本の神経が切断されると,それに伴っておこる現象はつぎの規則によっている.すなわち“それが属する神経細胞との結合が絶たれた線維が変性する.それにひきつづいておこる再生では,神経細胞と結合している中心端の軸索がのびて末梢端の変性した束の中に入り,この束の経路にしたがってそれが進んでゆくのである”(ウォラーの法則Wallersches Gesetz).
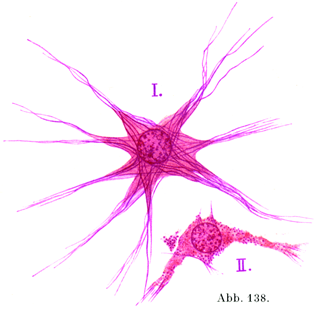
[図138]線維性グリア細胞 ヒトの脊髄より.グリア線維は紫. I. 線維の多いグリア細胞,II. 線維の少ないグリア細胞.
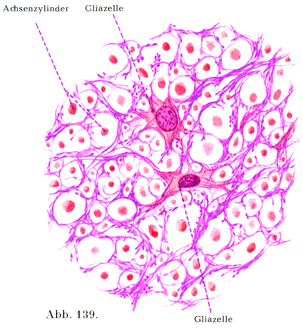
[図139]神経膠の網材 ヒトの脊髄の後索.
変性と再生の経過についてBetheが概観的に記載している.彼がいうところによれば,切断された神経の末梢株においてまず軸索が壊れて少数の片に分かれ,変性が進むにつれてこれが全く消えてします.ついには軸索は何も残らないまでにいたる.髄鞘もまた壊れて断片となり,これがますます短いものになって,終わりに全く(おそらく吸収されて)なくなる.変性の速さはここの動物部門によって異なる.かくしてシュワン鞘とその核と核をとりまく原形質だけが残っている.変性をあらわす変性は傷つけられた場所ではじまって,中心株の方も末梢株の方もそれが起きて波及する.そして末梢株は完全に変性してしまうが,中心株はある距離だけ死んで,その経過がそこで中止となる.変性の過程を条件付けるのは神経細胞とのつながりが切られたことではなくて,神経の傷つけられた個所の局所的障害が原因をなすのである.神経細胞じしんもまたその神経突起が切られたときは傷害を受けるのであって,細胞体内のニッスル小体が消えて,核は辺縁の方により,やはり傷害の徴候を示すのである.このできごとによって神経細胞が完全に死んでしまうことも少なくない.他の場合には細胞が後に回復する.そのとき核はふたたび中央部に戻って,ニッスル小体が新たにできる.
それゆえ切断された神経線維がふたたびくっついて治ったかどうかは初めのうちはどちらでもよいのである.しかしその合一がいつまでも失われていると,そのもとをなす神経細胞何ヶ月あるいは幾年かの内に萎縮してゆく.
若い動物では神経細胞から全く切り離され末梢株の中で再生がおこるのである(自発的再生autogene Regeneration, PhilippeauxとVulpian, Bethe).そのときは第4日頃からシュワン鞘の核と原形質がさかんな増殖をはじめる.20日間たつと変性した髄質の残りがほとんど全くなくなる.シュワン鞘の核と原形質が帯のような線維すなわちBandfasern(帯線維の意)をつくる.その中で外方の被う部分と中軸をなす部分が分かれてきて,後者はすでに原線維性のすじのようなものが存在する.その後に髄質の被いができる.それも核の近くでまずあらわれるのである.このさいなおわかることは,この再生現象は末梢株の中心端のところで末梢の部分よりもいっそう速やかに進行すると,中心部に近いほどそれが大きいということである.しかし自発的再生によってできた神経はのちにふたたび変性してしまう.
成長した動物でも自発的再生という現象は起きるが,それは中絶してします.つまり神経線維に自らその再生を完了するだけの力がない.再生をやり通すことは神経の中心端の影響があって初めて可能である.中心端は末梢株の仕事似た強いてただそれを激励するだけのはたらきをもつのである.Bethe, Folia neurobilogica 1907. この問題のいっそう新しい総括はBoekeの論文(D. Z. Nervenheikunde,115. Bd.,1930)にある.
中枢機関の中にある神経組織が傷ついたり物質欠如をおこした場合に,いかような変性および再生の現象がおこるかという問題は今後の研究に待たなければならない.残っている神経細胞が増殖することによって神経物質が新たに作られことは成長した温血動物ではおそらく全然おこらないのである.しかし硬骨魚類ではそれがKirsche, W. (Die regenerativen Vorgänge am Rückenmark erwachsener Teleostier nach operativer Kontinuitätstrennung. Arch. mikr.-anat. Forsch., 50. Bd.,1950)によって証せられた.同じ人の論文“Uber Neubildung der Gangllienzellen・・・”がVerh. anat. Ges.,1950に載っている.
化学的検査の結果,神経細胞がだいたいにおいて発生可能な細胞の組成を持つこと,しかしレチチンとコレステリンを比較的多く含むことに特色があることが分かった.なお成長した体の神経細胞の核にはヌクレインがわずか痕跡的にあるか,胎児の神経細胞にはこれが豊富である.
神経諸器官の二次的な成分としては次ぎにあげるようないろいろの物質がある::ノイロケラチン,ケファリン,プロタゴン,クレアチン,タウリン,グリコーゲン,乳酸,イノシット,ナトリウム化合物.尿素とロイチンの存在は疑問である(Kossel).
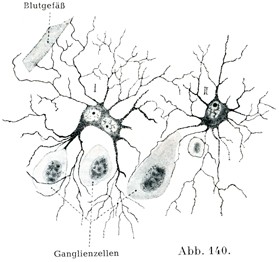
[図140]原形質性のグリア細胞 ヒトの大脳皮質より.(K. Schaffer,1915)Iは2核をもつ細胞:IIは1核をもつ細胞.
成人における細胞の総数Zahl der Zellen des Erwachsenen
成人の平均体積は600000ccm
1個の細胞の平均体積0.000008ccm
細胞の全数(血液を除いて)はおよそ8兆.
この章の本ですでにあげた文献の他に,次に述べる教科書は組織学を全般的にとりあつかったものである.
Bargmann, W., Histologie und mikroskopische Anatomie des Menschen. G. Thieme, Stuttgart, Bd. I,1948, Bd. II,1951.-Gegenraur-Furbringer, Lehrbuch der Anatomie d. Menschen. Bd. I, Viii. Aufl. Leipzig.1909.-Herrath, E. von und Abramow, S., Atlas der normalen Histologie und mikroskop. Anatomie des Mensche. G. Thieme, Stuttgart ,1950.
-Krause, R., Enzyllopaedie der mikr. Technik. Bd. I-III. Berlin,1926~1927.-Möllendorff, W. von, Handbuch der mikroskopischen Anatomie des Menschen. Berlin,1927-52.-Oppel, A., Lehrbuch der vergleichenden mikroskopischen Anatomie. Jena.1896, -Ranvier, L., Traité technique d’histologie. Paris,1888.-Schäfer, E. A., Text-Book of Microscopic Anatomy. London,1912.-Schaffer, J., Lehrbuch der Histologie. 3. Aufl.1933.-Sobatta, Lehrbuch und Atlas der Histologie usw. V. Aufl. München,1938.-Stöhr, Ph., Lehrbuch der Histologie. Jena. 26. Aufl. von W. v. Mollendorff.1949.-Stöhr, Ph. jr., Lehrbuch der Histologie und der mikroskopischen Anatomie des Menschen. Berlin,1952.
1-05

最終更新日 13/02/03